Overview cell membrane (Membrana
cellularis):
Pages with explanations are linked to the
text below the images if available! (Labelling is in German)
 |

|
 |
 |
 |
 |
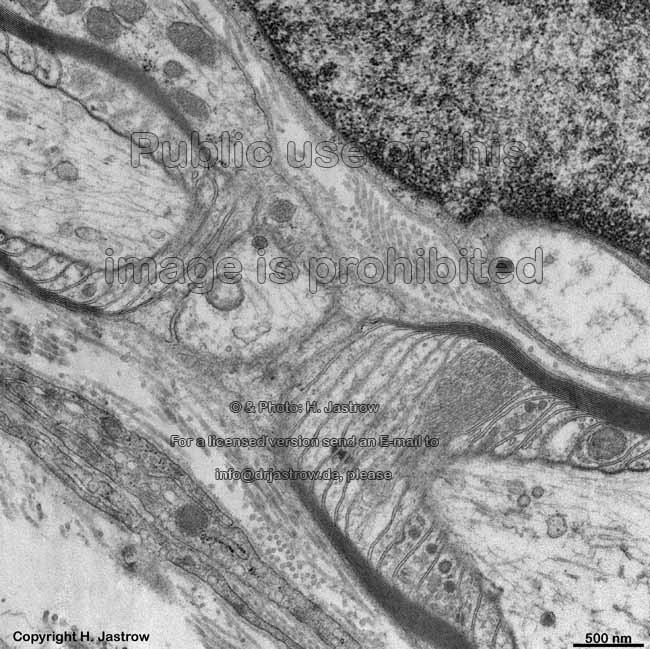
2 cell membranes and
intercellular space (rat) |
freeze fracture image of a
human erythrocyte membrane |
stacks of membranes of an
outer segemnet of a rod (rat) |
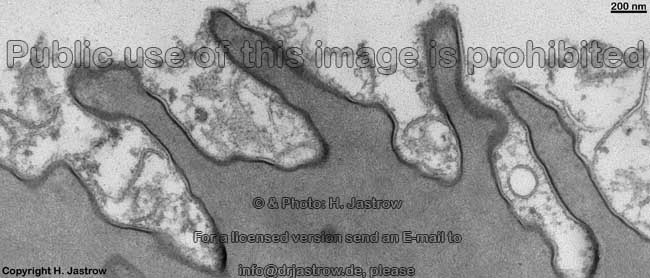
infolding of the membrane
in endocytosis (rat) |
thickened outer layer of the cell
membrane in urinary bladder (rat) |
cell membrane of
an erythrocyte (rat) |

|
 |
 |
 |
 |
 |
 |
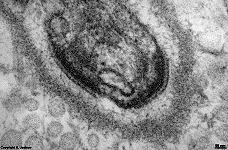
cell membrane of a
human kinocilium |
microvilli with glycocalix coat on
a human cell membrane |
protrusion of the cell membrane
in form of a microvillus (human) |
human microvilli in
cross section |
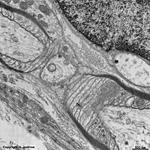
digitations = interdigitated
cell membranes (rat) |
myelin sheath consisting of multiple
layers of cellular membrane (rat) |
cell membrane at
human node of Ranvier |

|

|

|
|

|

|

|
ureter: formation of
membrane vesicles |
such vesicles in a super-
ficial cell of pig ureter |
irregular surface of
superficial cell (pig) |
plaque attached to the cell mem-
brane in a hemidesmosome (rat) |
stable connection of cells involving cell
membrane: desmosomes, human skin |
membrane vesicle
formation ureter (pig) |
detail: thickened outer layer
of the cell membrane ureter (pig) |
 |
 |
 |
 |
 |
 |
disruption of the membrane during
secretion of a goblet cell (rat) |
neurotransmission in a synapse
happens on the cell membrane (rat) |
receptors in cell membrane
are required for olfaction (rat) |
tight junction of cell membranes
as
basis for blood liquor barrier
(rat) |
keratinized squamous epithelial cell with thickened
cell membrane of esophagus (rat) |
very closely attached cell mem-
branes with channels in a nexus (rat) |
The cell membrane (Terminologia histologica: Plasmalemma;
Membrana cellularis) also called plasmalemma or cytomembrane
is a biological unit membrane, i.e. is a double-membrane.
It constitutes the outer border of all cells
in humans and animals limiting the (interior) cellular space, i.e. the
cytoplasm
or cellular body with its organells which is also called intracellular
space to the surrounding extracellular space. The cell membrane is too
small to be visible with a light microscope due to its thickness of only
6
to 9 (mostly 8) nm. Nevertheless this small elastic "skin" of the cell
is rather solid.
Biochemical structure:
As biological unit membrane the cell membrane consists of lipids
and proteins which may be connected to sugars and linked to its
surface as glycolipids or glycoproteins to form an additional layer, the
glycocalyx.
The relation of lipids to proteins ranges from 4 : 1 to 1 : 4 and depends
on the kind of the cell and its metabolic activity. Most of the lipids
involved in cell membrane construction are phospholipids which are
further classified as glycerophosphatids (phosphatidylethanolamin, phosphatidylcholin,
phosphatidylserin) and sphingophosphatids (sphingomyelin, cerebrosids,
gangliosids). Additionally cholesterin is present as neutral lipid.
The outer part of the cellular membrane mainly shows glycoproteins and
glycolipids or sphingomyelin and phosphatidylcholine whereas the inner
membrane has a highe amount of phosphatidylethanolamin; phosphatidylserin
is only seen in the inner layer. Most of the lipids have a polarhead
part which is hydrophilic, i.e. has a high affinity for water while
being rejectant to lipids, i.e. lipophobic. The apolar tail part of lipids
is formed by 2 long fatty acid chains (one of which often is insaturated)
and is rejectant to water, i.e. hydrophobic but lipophilic, i.e.
attacting lipids. Thus the whole lipid has an amphitathic character meaning
that both molecular ends show opposing affinities. When a larger amount
of such molecules comes into a waterly medium as it is present in the body,
they arrange to spherical bodies spontanously. These spheres are partly
micells and partly liposomes. Liposomes are spherical arrangements
of amphipathic lipids in which the hydrophilic head parts are directed
outwards while the ends of the tails are oriented to the interior.
Micells
are also spherical arrangements but of a double membrane that encloses
a little fluid. Their composition is as follows: hydrophilic head parts
are directed outwards side by side, in the mid region the lipophilic tail
ends of the two layers are touching each other and the hydrophilic heads
of the 2nd membrane layer are oriented to the interior which contains waterly
medium;
briefly: outwards - heads
of lipid 1 - tail of lipid 1
- tail of lipid 2 - head
of lipid 2 - inner space of the micell.
The cell membrane is similar to a micell but further has a lot of different
associated or integrated proteins. The latter are either peripherical
or integral. Peripherical proteins are electrostatically bound to
the outer polar heads of the lipids of the cell membrane, thus they are
either attached to the outer surface or the cytoplasmic surface of the
membrane. Integral proteins reach with hydrophipic molecule areas
into the hydrophibic central area of the double-membrane. Dependent on
their molecular composition large proteins may reach through the entire
membrane and show parts on the outer as well as inner membrane surface.
Such proteins are called transmembrane proteins. In many cases such
proteins form a minute pore in their interior which connects the extra-
to the intracellular space allowing entry of water or ions into the cell
or the contrary, i.e. allow water / ions to leave the cell. Such ion
channels are of special importance for exicitatory processes and conductance
e.g. in smooth or heart
muscle cells. The opening and closure of all such tunnel proteins
is well controlled, mostly specific for one or two kinds of ions and combined
with a change of their three-dimensional structure. In some cases the transported
substrate alone e.g., glucose after binding to a specific extramembrane
area of the protein opens the channel in other cases a further (mostly
hormoneal) signal is required for channel activation.
All components of the cellular membrane are free free floating,
i.e. may move freely in any direction resulting in a fluid mosaic membrane
model. The movement increases with temperature. At low temperatures
cell membranes are thinner (6 nm) and have a gel-like consistency
whereas at high temperatures membrane thickness may raise to 9
nm and the consistency gets sol-like. At normal body temperature
cellular membranes are ~8 nm thick and sol-like.
Electron microscopic composition:
Electron microscopic examination shows 3 layers of the cell
membrane:
- outwards, i.e. at the outer or
superficial surface (Terminologia histologica: Facies externa) a 2.5
nm thick external electron-dense lamina (Terminologia histologica:
Lamina densa externa) consisting of the hydrophilc portions of the lipids
with integrated or attached proteins. At some cells a glycocalyx
(Terminologia histologica: Glycocalyx) consisting of sugars and attached
proteins is anchored to the external cellular membrane and reaches into
the extracellular space. In freeze-etching preparations where a specimen
is broken by special aparatuses this outer face of the cell membrane is
called E-face, external fractured face or exoplasmic face (Terminologia
histologica: Facies E; Facies fracta externa).
- in the centre a 3 nm strong hardly
electron-dense layer, the middle lucent lamina (Terminologia histologica:
Lamina intermedia lucida) in which the hydrophobic ends of the lipids and
hydrophobic regions of transmembrane proteins are located. Rarely intramembrane
particles, i.e. fine electron-dense granules (Terminologia histologica:
Granula intramembranacea), protrusions of intramembrane particles (Terminologia
histologica: Protrusiones granulorum intramembranaceorum) or rare impressions
of intramembrane particles (Terminologia histologica: Impressiones granulorum
intramembranaceorm) are seen in this area.
- the inner lamina inner surface
or cytosolic face which is directly bordering the cytoplasm
(Terminologia histologica: Facies interna) is a 2.5 nm thick
internal electron-dense lamina (Terminologia histologica: Lamina
densa interna). Filaments of the cytoskeleton
are anchored to the proteins of this lamina. In freeze-etching preparations
this face of the cell membrane is called protoplasmic fractured face; P
face (Terminologia histologica: Facies P; Facies fracta cytoplasmica; Facies
fracta protoplasmica).
One image above shows such a freeze etching preparation. Hereby the
double membrane is broken in the central area where the tails of the lipids
hit each other, thus the outer (exoplasmatic = E-face) and the inner (protoplasmatic
= P-face) get apparent. Using this technique the broken transmembrane proteins
and fine regular tails of the lipids may be differentiated.
Function
- The cell membrane is a border between the extra- and the intracellular
space which show considerable differences in composition. All substances
that enter or leave cells have to pass or need to be transported throgh
it. Cell membranes are semipermeabel, i.e. allow only few substances
with certain chemical behaviour to pass. Small lipophilic molecules e.g.,
steroid hormones and hormones of the thyroid
gland may directly pass the plasmalemm, whereas all larger or hydrophilic
substances require special transport mechanisms which usually base on more
or less specific proteins that mostly require energy for such active transport
processes.
- The shape of cell membranes follows changes of the cytoskeleton
and plastically adapts to them e.g., when processes or pseudopods
are newly formed or redrawn into the soma in freely moving cells. Cell
membrane material may be transported throgh the whole cell to an opposite
side via vesicles.
- electric impulses spread along the cell membrane which is
of major importance in nerve cells and their processes.
- The glycocalyx which is present
on most cells is very important for cell recognition by the immune system
and the morphological substate of blood groups.
- The integral proteins of the inner lamina of the cell membrane bind
to filaments of the cytoskeleton
to preserve stability of the whole cell.
- The functions of cells are influenced by extracellular signals which
are the targets of lots of receptors mostly located on the surface
of the cell membrane.
In fact most of the membrane bound proteins serve as receptors
for mostly highly specific substrates (ligands). This means that
only very few substances with very special chemical and structural components
are able to interact with the receptor molecules. There are many classes
of receptors e.g., receptors for cellular growth and differentiation, receptors
for neurotransmitters (of nerve cells), immunological receptors for cell
recognition, for viruses or bacterial toxins, receptors to which pharmaca
bind. On the cytoplasmatic face of the cell membrane receptors with affinity
to cytoplasmatic filaments predominate.
The binding of a ligand to its receptor results in a structural change
of the latter which causes different kinds of effects:
A. signal transduction:
The three-dimensional structure of the receptor protein is altered by the
specific interaction. Three different kinds of such changes are known:
Type 1 proteinphosphorylation with tyrosinkinase: after
binding of the ligand the changed transmembrane receptor activates another
protein at the cytoplasmic side under ATP consumption e.g., the insulin
receptor;
Type 2 ligand activated ion channel e.g., a neurotransmitter
like acetylcholin opens a sodium channel at a synapse;
Type 3 G-protein mediated release of a second messenger
like e.g., cyclic Adenosinmonophosphate (c-AMP), Innositoltriphosphate
(IP3) or cyclic Guanosinmonophosphate (c-GMP), in these cases intermolecular
processes with neighbouring proteins cause activation of an enzyme located
on the cytoplasmic surface of the plasmalemm.
B. receptor induced Endocytosis:
At this occasion a small invagination of the cell membrane allows intake
of extracellular fluid and molecules by formation of a vesicle which further
migrates into the cytoplasm.
cell membrane junctions:
In epithelia cells are connected to each
other via different intercellular junctions to ensure tissue stability.
The cell membrane is involved in formation of such cell-to-cell
connections. Besides interdigitations
of cell membranes the following special contacts occur: Zonula
occludens and adhaerens, Fascia
adhaerens as well as Nexus and Desmosome.
surface differentiations of cells:
Cells can effectively raise their surface area by protrusions
of their membranes which can be seen in resorbing epithelia.
Depending on morphology and function plasmalemma-coated cell protrusions
like immotile microvilli or stereocila
can be differentiated from mobile processes like kinocilia,
cilia
and pseudopods. The uppermost coating cells
of transitional epitelia e.g., ureter or urinary
bladder, show a very electron-dense and thick outer lamina of their
surface membrane towards the lumen. At synapses
pre- and postsynaptic densities are attached to the cell membrane. In smooth
muscle cells, endothelial cells and
hair
cells small round dips of the extracellular space into the cytoplasm,
the caveols are typical. In rods and cones
of the retina dense membrane stacks deriving
from the plasmalemm bind the visual pigments.
--> cell surface specialisations, microvilli,
kinocilia,
glycocalix,
cell-to-cell
contacts, synapse, gap
junction, tight junction
--> Electron microscopic atlas Overview
--> Homepage of the workshop
One image was kindly provided by HSD Dr. Klinger;
other images, page & copyright H. Jastrow.