| [Все] [А] [Б] [В] [Г] [Д] [Е] [Ж] [З] [И] [Й] [К] [Л] [М] [Н] [О] [П] [Р] [С] [Т] [У] [Ф] [Х] [Ц] [Ч] [Ш] [Щ] [Э] [Ю] [Я] [Прочее] | [Рекомендации сообщества] [Книжный торрент] |
Как мы видим? Нейробиология зрительного восприятия (fb2)
 - Как мы видим? Нейробиология зрительного восприятия (пер. Ирина Вадимовна Евстигнеева) 2192K скачать: (fb2) - (epub) - (mobi) - Ричард Маслэнд
- Как мы видим? Нейробиология зрительного восприятия (пер. Ирина Вадимовна Евстигнеева) 2192K скачать: (fb2) - (epub) - (mobi) - Ричард МаслэндРичард Маслэнд
Как мы видим? Нейробиология зрительного восприятия
Переводчик Ирина Евстигнеева
Научный редактор Оксана Агеенкова
Редактор Любовь Любавина
Главный редактор С. Турко
Руководитель проекта А. Василенко
Корректоры Е. Чудинова, Е. Аксенова
Компьютерная верстка А. Абрамов
Дизайн обложки А. Бондаренко
Иллюстрация на обложке Shutterstock.com
© 2020 by Richard Masland
© Издание на русском языке, перевод, оформление. ООО «Альпина Паблишер», 2022
Все права защищены. Данная электронная книга предназначена исключительно для частного использования в личных (некоммерческих) целях. Электронная книга, ее части, фрагменты и элементы, включая текст, изображения и иное, не подлежат копированию и любому другому использованию без разрешения правообладателя. В частности, запрещено такое использование, в результате которого электронная книга, ее часть, фрагмент или элемент станут доступными ограниченному или неопределенному кругу лиц, в том числе посредством сети интернет, независимо от того, будет предоставляться доступ за плату или безвозмездно.
Копирование, воспроизведение и иное использование электронной книги, ее частей, фрагментов и элементов, выходящее за пределы частного использования в личных (некоммерческих) целях, без согласия правообладателя является незаконным и влечет уголовную, административную и гражданскую ответственность.
* * *
Введение
Это книга о том, как мы видим. Мыслители издавна пытались объяснить феномен зрения, но их представления, как показывают современные исследования, были наивными: ведь глаз – это не просто съемочный аппарат, а нечто гораздо большее. Наша способность узнавать лица друзей кажется элементарной и естественной – настолько, что древние даже не рассматривали ее как предмет исследования, – но на деле в ней нет ничего очевидного. Чтобы дать исчерпывающий ответ на вопрос, что такое зрение, надо понять не только то, как функционируют наши глаза. Необходимо также знать, как наш мозг воспринимает и осмысляет внешний мир.
Как это ни парадоксально, мозг очень медлителен: нейроны и синаптические связи между ними функционируют в миллионы раз медленнее современных компьютеров. Однако он превосходит компьютеры в большинстве перцептивных задач. Мы можем за тысячные доли секунды узнать своего ребенка в толпе мальчиков и девочек на детской площадке. Как наш мозг это делает? Каким образом он перерабатывает и осмысляет потоки элементарных стимулов – пятен света, вибраций воздуха, давлений на кожу и т. п.? На сегодняшний день у нас есть только проблески понимания, но от того, что уже известно, захватывает дух.
Я пришел в нейронауку в 25 лет – еще до того, как она стала самостоятельной официальной дисциплиной, – и сегодня увлечен ею так же страстно, как и тогда. Я наблюдал за развитием нейронауки и принимал в нем непосредственное участие. Эту книгу я написал, чтобы рассказать вам о том, как работает зрение – от сетчатки до зрительных центров в височной коре мозга. Но еще я хочу пригласить вас в научное путешествие, чтобы вы узнали, как работают нейробиологи, не из скучных научных статей или ток-шоу, а увидели это своими глазами – побывав в настоящих исследовательских лабораториях. Наконец, я познакомлю вас с несколькими ключевыми фигурами в этой области.
Мы шаг за шагом рассмотрим процесс зрительного восприятия. Вы узнаете, что мы видим мир вовсе не таким, какой он есть на самом деле: наша сетчатка разбивает его на множество отдельных фрагментов (сигналов) и посылает их в мозг по отдельным каналам, каждый из которых несет информацию об одном небольшом аспекте изображения. Я объясню, как нейроны сетчатки выполняют это перекодирование и почему. Затем мы проследуем за этими сигналами в мозг и посмотрим, как из них формируется восприятие.
В мозге по-прежнему еще множество тайн, но мы уже смогли прийти к важному выводу: бо́льшая часть мозга работает не как система фиксированных двухточечных соединений наподобие телефонной сети, а как паутина бесчисленных нейронных связей, то есть как нейронная сеть. В наши дни нейронные сети обычно ассоциируются с компьютерами, но их идея впервые была выдвинута более полувека назад прозорливым канадским нейробиологом Дональдом Хеббом. Несколько лет спустя эту теорию подхватили специалисты в области теории вычислительных систем. В последующие десятилетия нейронные сети то входили в моду, то теряли популярность, но более совершенные компьютеры в конечном итоге привели к рождению новой области искусственного интеллекта (ИИ), известной как машинное обучение. Разработчики ИИ показали, что компьютерные нейронные сети могут научиться впечатляющим вещам, и тем самым побудили нейробиологов вновь посмотреть на головной мозг сквозь призму нейронных сетей. Сегодня у нас есть замечательный альянс нейробиологии и компьютерных наук, в котором каждая дисциплина служит источником идей для другой.
Действительно ли мозг использует нейронные сети для восприятия и осмысления мира? Функционирует ли он согласно принципам, применяющимся в «машинном обучении»? Ответ, судя по всему, да – и мозг делает это намного лучше компьютеров. Безусловно, компьютеры поражают нас некоторыми своими способностями – не только игрой в шахматы, но и выполнением других более сложных задач. Но по большому счету они как цирковые пони, умеющие делать только один трюк. И даже самые простые системы ИИ требуют большого количества оборудования и, как следствие, большого количества энергии. В отличие от них, наш скромный по размерам мозг способен выполнять огромное разнообразие задач, потребляя при этом меньше энергии, чем ночник для чтения. С этой точки зрения компьютеры очень примитивны, поэтому цель – сделать их хотя бы немного похожими на человеческий мозг.
Как это давно понял Дональд Хебб, нейронная сеть с фиксированными соединениями неспособна учиться. Ключ к обучаемости нейронной сети (биологической или искусственной) – в способности синаптических связей между ее нейронами меняться под влиянием опыта. Такая пластичность – общее правило для всего мозга, а не только для сенсорных систем. Благодаря ей мозг может оправляться от повреждений и выделять дополнительные ресурсы под особенно важные задачи. В зрительной системе нейронные сети могут научиться заранее идентифицировать визуальный объект, дополняя сенсорную информацию, поступающую с сетчатки, знаниями об увиденных ранее аналогичных объектах. Это означает, что значительная часть нашего восприятия – не столько фиксированная, сколько приобретенная в результате обучения реакция на зримый объект. Нейронные сети распознают определенные комбинации признаков, когда они их видят.
Куда это приведет нас в наших поисках понимания механизмов восприятия, мышления, эмоций? Конкретные детали нам неизвестны, но мы можем заглянуть в далекое будущее и попробовать увидеть ответ. Это будут фактологические, поддающиеся проверке научные знания в каждой точке. В этой книге мы с вами пройдем часть пути – до того места, где сенсорный опыт превращается в восприятие и мысль.
Наконец, где во всем этом наше «я»? Легко говорить о мозге с позиции внешнего наблюдателя, но что представляет собой – и где обитает – тот «внутренний человек», который смотрит на мир нашими глазами? Задумываясь об этом, мы неизбежно наталкиваемся на вопрос о природе сознания, нашего «я», а на этом пути мы, люди, едва ли сделали первые шаги. Я затрону эту тему в конце книги, не давая ответов, но постаравшись чуть более четко обозначить проблему.
Часть I
Зрение: Первые шаги к пониманию
В 1960-х гг. в Гарвардском университете уважаемый профессор Джейкоб Бек читал курс, незатейливо назвавшийся «Восприятие». Лекции проходили в небольшой аудитории, втиснутой в углу Мемориального холла – величественного готического здания из темно-красного кирпича, возведенного в XIX в. в память гарвардцев, погибших в Гражданской войне. Около сотни коричневых деревянных столов, покрытых за прошедшее столетие бесчисленными слоями пожелтевшего лака, ступенчато спускались к огромной черной доске, которая занимала всю переднюю стену. Через редкие окна, расположенные высоко на левой стене, виднелось холодное небо, но лампы накаливания заливали теплым желтым светом аудиторию, в которой находилось 30–40 студентов.
Манера преподавания Бека была такой же простой и ясной, как и название его курса. Он придерживался классического стиля, считая, что его главная задача – четко и организованно изложить учебный материал, а не увлечь студентов. В ходе лекций он пользовался тщательно подготовленными конспектами и в начале каждого занятия обязательно уделял несколько минут тому, чтобы повторить основные моменты предыдущей лекции.
Впрочем, Беку и не надо было вести себя как шоумен. Материал захватывал сам по себе. Разумеется, Бек преподавал нам основы: давление на кожу деформирует нервные окончания, которые посылают сигнал через спинной мозг в головной. Одни наши кожные рецепторы реагируют на прикосновения, другие – на тепло, третьи – на движение по коже, например, когда вы гуляете по лесу и вдруг чувствуете, что у вас по руке ползет неизвестное (и, возможно, ядовитое) насекомое. Такого рода факты любого могли заинтересовать. Но самой удивительной проблемой, которой Бек озадачил нас, 19-летних, была проблема распознавания объектов.
С одной стороны, она непосредственно связана со зрительным восприятием: как работает глаз, как он подает сигнал в мозг. Но не только. Здесь играют роль куда более сложные феномены, такие как мышление, память, природа сознания. Сегодня мы более-менее представляем себе, как работает наша сенсорная система. Мы научились регистрировать прохождение электрических сигналов по чувствительным проводящим путям. Научились стимулировать нейроны, что позволяет нам все больше узнавать об их работе и функциях. Мы многое узнали о том, как обрабатываются сенсорные сигналы в головном мозге и как они передаются между различными его отделами. Таким образом, у нас появились надежные базовые знания, опираясь на которые мы можем двигаться дальше по пути, на котором мы только-только начинаем делать первые шаги, – по пути к пониманию того, как мыслит наш мозг. И изучение феномена нашего зрения обещает по крайней мере частично приподнять завесу над великими тайнами.
1 | Чудо восприятия
Груши – не скрипки,Не обнаженные тела, не бутыли.Они ни на что не похожи.Они – желтые формы,Сотворенные из изгибов,Бочковатые книзу,Чуть тронутые красным.УОЛЛЕС СТИВЕНС
Взгляните на эти три лица. Хотя фотографии несколько размыты и неконтрастны, вы легко можете распознать, кто на них изображен: справа – женщина (у нее более округлое лицо); в центре – мальчик (у него явно мужской подбородок). Будь они вашими сыном или дочерью, братом или матерью, другом или подругой или любым другим близким человеком, вы бы узнали их в любом виде и в любой ситуации, в профиль и анфас, при ярком свете и в сумерках, вблизи и издалека, радостными, грустными, смеющимися или молчаливыми.

Задумывались ли вы когда-нибудь о том, как вы это делаете? Каждый раз на вашу сетчатку падают фактически разные изображения. Ваш мозг приспосабливается к каждому изображению – крупному либо мелкому, яркому либо тусклому, с улыбкой или нахмуренными бровями. Число различных версий лица – как физического раздражителя, воспринимаемого вашей сетчаткой, – практически бесконечно. Однако же мы узнаем знакомые лица мгновенно и без усилий. И мы способны различать не три лица, а сотни и тысячи. Каким же образом нашему мозгу, который, по сути, является всего лишь природным аппаратом, как и все остальное в нашем теле, удается так хорошо справляться с этой задачей?
Давайте начнем с более простого примера. Представьте, что вам нужно разработать компьютерную программу, способную распознавать букву А. Современные компьютеры справляются с этим легко, не так ли? Но это всего лишь видимость – компьютеры нас обманывают (через пару абзацев я объясню, почему так говорю).
Решение кажется очевидным: в компьютере (или в мозге) должен иметься шаблон или образец буквы А. Компьютер (или мозг) сравнивает распознаваемую букву с образцом буквы А и делает вывод об их сходстве или различии. Но что, если размер распознаваемой А отличается от размера шаблонной А? Их сопоставление покажет, что это разные буквы.
Хорошо, значит, в компьютерную программу необходимо включить все множество шаблонов буквы А разного размера:

Окей, проблема с несхожими размерами решена. Но предположим, что распознаваемая буква А немного наклонена влево:  Накренившаяся буква снова не будет совпадать ни с одним из имеющихся шаблонов.
Накренившаяся буква снова не будет совпадать ни с одним из имеющихся шаблонов.
Чтобы решить эту проблему, мы включаем в программу набор шаблонов буквы А всех возможных размеров со всеми возможными углами наклона. Если компьютер достаточно мощный, эта программа может работать достаточно быстро. Но как насчет остальных параметров, таких как толщина линии, цвет, шрифт и т. д.? Мало того: число комбинаций, которые в итоге нужно проверить компьютеру, – все возможные размеры, умноженные на все возможные углы наклона, умноженные на все возможные свойства шрифта, умноженные на все возможные цвета, и т. д. Это количество становится очень большим, слишком большим с практической точки зрения. И вся морока ради того, чтобы распознать одну-единственную букву!
С лицами числу вариантов и вовсе нет предела. Лицо может улыбаться или хмуриться, быть ярко освещенным или находиться в тени, быть видимым в профиль или анфас. А элементы нашего мозга – нейроны и синапсы – работают очень неспешно по сравнению с современными компьютерами. Передача самого базового сигнала через синапс между контактирующими нейронами занимает около тысячной доли секунды. За это время мощный компьютер успевает выполнить почти миллион операций. Именно благодаря такой сверхчеловеческой скорости компьютеры и могут обманывать нас, делая то, на что неспособен ни один живой биологический организм. Если предположить, что сравнение по одному из параметров состоит из ста операций, то, пока наш мозг передает через синапс один нервный импульс, компьютер успевает сравнить тысячу параметров. И это без учета времени прохождения сигнала по соединяющим нейроны нервным волокнам (аксонам)! Если бы наш старина мозг работал по тому же принципу, что и компьютеры, ему требовались бы минуты, чтобы распознать даже хорошо знакомое лицо. Короче говоря, перебор вариантов – не вариант для нашего мозга.
Вот пример, связанный с другим восприятием – слуховым[1]. Возьмем так называемый феномен сегментации. Если кто-то вам скажет: «Вон бежит синяя собака» – вы услышите то, что будет примерно соответствовать написанному выше. Но на самом деле в обычной устной речи мы не делаем пауз между словами (не считая тех случаев, когда мы выделяем каждое слово намеренно). С акустической точки зрения вы слышите эту фразу как один непрерывный звуковой поток: «Вонбежитсиняясобака». Чтобы осмыслить ее, наш мозг разбивает эту длинную последовательность звуков на отдельные знакомые нам слова.
И снова очевидно, что мозг делает это не путем сопоставления слов с некими шаблонами. Сколько звуков должны были бы включать такие шаблоны? Сколько словесных форм? Разумеется, гораздо больше, чем в словаре. И это не говоря уже о разных акцентах, темпах речи, фоновом шуме и многом другом…
Эта загадка – способность, которую мы с такой легкостью используем по многу раз на день, – и есть то, что мы называем проблемой распознавания объектов. Хотя ее принято рассматривать в основном как проблему восприятия, здесь также задействована память: чтобы распознать объект, нам нужно сопоставить текущий раздражитель с воспоминаниями о соответствующих объектах, с которыми мы сталкивались в прошлом. Выяснить, как это работает, – захватывающая научная задача, Эверест сенсорной нейробиологии.
2 | Нейроны, рассказывающие мозгу о внешнем мире
Мы изучаем общее
через изучение конкретного.
СТИВЕН КУФФЛЕР
Как я уже предупредил вас, мир, который мы видим, – вовсе не тот мир, что существует на самом деле. Наша сетчатка анализирует воспринимаемое визуальное изображение, выделяет в нем наиболее значимые компоненты, какие-то из них модифицирует и посылает десятки отдельных потоков сигналов о каждом из них в наш мозг, который собирает из них «видимую» нами картину миру. Все остальное рассматривается как фоновый шум и игнорируется. Такое упрощение сенсорной сигнализации – не просто эволюционная прихоть, а один из наиболее фундаментальных принципов всего восприятия, главная цель которого – экономия.
Чтобы понять, как это работает, давайте начнем с основ.
НЕЙРОН
Нейрон – штука довольно простая. Это крошечный физический объект, состав которого нам понятен. Он включает в себя те же компоненты, которые входят в любую животную клетку, но которые, однако, имеют ряд уникальных особенностей. Когда несколько сотен миллионов нейронов объединяются в сеть, происходят поистине фантастические вещи: мы, владельцы этой нейронной сети, можем узнавать друзей, наслаждаться музыкой Бетховена или ловить мяч одной рукой с расстояния 27 м.

Нейрон, как и все клетки позвоночных, представляет собой мешочек с внутриклеточной жидкостью, отделенный от окружающей среды тонкой эластической мембраной. Одни нейроны похожи на детские воздушные шарики. Форма других более сложна: они походят на амеб. Третьи и вовсе поражают своим причудливым строением. Большинство нейронов напоминают голые деревья зимой с многочисленными ветвями и веточками – с помощью этих отростков они соединяются с другими нейронами, своими ближайшими и дальними соседями. Но при всей замысловатости форм нейрон, как и любая другая клетка, состоит из единого внутреннего пространства, заключенного в границы мембраны – даже если местами эта мембрана напоминает не привычный мыльный пузырь, а тончайшие изогнутые трубочки для напитков.
Что же представляет собой клеточная мембрана? Она состоит из липидов – разновидности жиров, которые, как известно, не смешиваются с водой. Благодаря этому мембрана и выполняет свою барьерную функцию между внутренней и внешней водными средами. Но сама по себе эта липидная оболочка мало что может делать. В лабораторных условиях можно создать искусственную клетку, имеющую одну только клеточную мембрану, но такая клетка будет лежать мертвым грузом. Настоящая клеточная мембрана усеяна мириадами крошечных молекулярных машин, каждая из которых выполняет свою конкретную задачу – например, один из видов встроенных в нее белковых молекул открывает «ворота» (каналы), пропуская внутрь клетки и из нее потоки заряженных частиц (ионов). Этот механизм лежит в основе передачи нервного импульса.
Нервные клетки выполняют множество функций, но главная из них – та, что отличает их практически от всех остальных клеток, – коммуникация с другими нейронами. В большинстве случаев они делают это посредством передачи коротких электрических импульсов, или спайков. Эти импульсы могут передаваться как на короткие, так и на очень длинные расстояния. Некоторые нейроны поддерживают коммуникацию (мы говорим: «Обмениваются нервными импульсами») только со своими ближайшими соседями. Эти так называемые интернейроны (вставочные нейроны или нейроны локальной сети) передают сигналы на расстояние до 10 микрометров, то есть всего до одной сотой миллиметра (1 мкм равен 0,001 мм). Для сравнения: некоторые нервные импульсы проходят путь от головного мозга до нижней части спинного мозга, когда вы пытаетесь пошевелить большим пальцем ноги, или в обратном направлении, если вы больно бьетесь ногой о лежащий на земле камень.
Нервные импульсы передаются через нейроны совсем не так, как электрический ток, текущий по медным проводам. Это гораздо более сложный биологический процесс, и в нем активно участвует клеточная мембрана: передача электрического импульса происходит за счет быстрого колебания мембранного потенциала, которое возникает в результате перемещения потоков ионов внутрь клетки и из нее через встроенные в мембрану специализированные белковые молекулы (ионные каналы). Вот почему передача импульсов происходит довольно медленно по сравнению с течением электрического тока. В зависимости от типа аксона скорость распространения по нему нервного импульса составляет от 10 до 100 м/сек, тогда как электричество бежит по проводам со скоростью около 300 млн м/сек. Такая низкая проводимость ограничивает вычислительную мощность нашего мозга и является основной причиной того, почему он не может использовать для решения проблем простые стратегии перебора, основанные на грубой вычислительной силе.
На конце аксона обычно находится синапс. Это место контакта, через которое нейроны общаются друг с другом. Возникший в синапсе нейрона электрический сигнал преобразуется в химический: под воздействием спайка специальный синаптический механизм выбрасывает в пространство между двумя синапсами (синаптическую щель) особые химические вещества, которые воспринимаются синапсом другого нейрона. Эти химические передатчики сигналов называются нейромедиаторами или трансмиттерами. Поскольку существует очень много разных типов нейромедиаторов, которые используются для разных целей в разных отделах мозга, а также благодаря тому, что механизм их выработки включает множество шагов, мы имеем возможность вмешиваться в этот процесс и в какой-то степени манипулировать функционированием мозга – с терапевтическими целями или ради удовольствия[2]. Например, мы можем воздействовать на синапсы с помощью нейролептиков, противоэпилептических препаратов, валиума, помогающего нам успокоиться, прозака, делающего нас счастливыми, а также хорошо известного всем никотина.
Выбрасываемые нейроном нейромедиаторы могут быть возбуждающими или тормозящими – могут повышать или понижать активность воспринимающего их нейрона. (В реальности нейрон редко подвергается воздействию только одного типа нейромедиаторов, но сейчас для простоты обсуждения давайте предположим, что это так.) Второй нейрон интегрирует все получаемые им входные сигналы, и, если за короткое время количество определенных сигналов превышает определенный порог, в нем возникает волна возбуждения – «потенциал действия». Этот потенциал действия распространяется по нейрону и передается дальше третьему нейрону, приводя к его возбуждению или торможению, и т. д.
На этом этапе мы видим вторую важную функцию нейронов: они решают, какие сигналы передать другим нейронам, а какие не пропустить дальше. Это решение принимается ими, как уже было сказано, путем объединения всех получаемых входных сигналов. Проще говоря, они складывают все возбуждающие сигналы и исключают из них все тормозящие. Разумеется, это очень упрощенная картина, учитывая все разнообразие входящих сигналов и факторов, влияющих на их действие. Изучением этого процесса занимается отдельная область нейробиологии: некоторые из умнейших моих коллег посвятили свою жизнь исследованию многочисленных и удивительных способов синаптической коммуникации.
Итак, базовая функция нейрона – ждать поступления химических входных сигналов и, когда эти сигналы достигают определенной величины, генерировать потенциал действия, чтобы передать нервный импульс. Но простая передача импульсов от нейрона к нейрону не делает мозг мозгом. Чтобы мозг был мозгом, необходимо, чтобы нейроны принимали решения о том, какие сигналы передавать дальше, а какие нет. Разумеется, я сильно упрощаю, потому что моя цель – рассказать вам о восприятии. А для этого вам нужно знать всего несколько ключевых вещей и прежде всего то, что потенциал действия вызывает изменение электрического заряда на своем пути и мы, простые смертные, можем отследить этот электрический импульс, или спайк, при помощи тончайших длинных зондов, называемых микроэлектродами.
КАК СЕНСОРНЫЕ НЕЙРОНЫ ПЕРЕДАЮТ СИГНАЛЫ
Как было сказано выше, нейроны могут передавать сигналы на короткие и очень длинные расстояния. Например, у жирафа нейроны, отвечающие за ходьбу и соединяющие головной мозг с нижним отделом спинного мозга, могут достигать в длину 2,5 м. Но во всех случаях (за редкими исключениями) способ передачи сигнала одинаков: воздействие раздражителя в каком-либо месте на поверхности клетки приводит к возникновению электрического импульса (потенциала действия), который распространяется по нейрону и передается дальше.
Все нейроны, отвечающие за восприятие внешнего мира – будь то через осязание, слух, зрение, вкус или обоняние, – по сути, делают одно и то же: они регистрируют событие и передают сигнал о нем – иногда через один-два вставочных нейрона – в головной мозг. Но совершают они это довольно разными способами, поскольку воспринимаемые ими внешние события также являются очень разными по своей физической природе.
Возьмем осязание. Тактильные ощущения возникают в результате деформации кожи под давлением. Эти деформирующие давления могут быть разными по силе – от удара о твердый предмет до легкого поглаживания (мы способны почувствовать даже невесомого комара, осторожно севшего нам на руку), – и все они воспринимаются нервными окончаниями, которые расположены под поверхностью кожи и являются частью нейронов.

На рисунке показаны два нейрона на проводящем пути тактильной чувствительности; участок кожи, обозначенный пунктирной линией, называется рецептивным полем (соответственно сенсорная информация на схеме перемещается слева направо). Первый нейрон имеет длинный отросток (аксон), идущий от кожи – где он разветвляется на множество тончайших нервных окончаний – к спинному мозгу. Когда вам на руку садится комар, его ноги слегка давят на кожу над нервным окончанием. Нейрон регистрирует это давление и генерирует нервный импульс. Импульс проходит по аксону через тело клетки и доходит до синапса (на рисунке обозначен раздвоенной линией), соединенного со вторым нейроном, который находится в спинном мозге и передает сигнал дальше в головной мозг. (Существуют и другие пути передачи сигналов в мозг. Это один из самых простых.)
Тактильные нервные окончания обнаруживают давление на поверхности кожи при помощи механочувствительных ионных каналов – специальных белковых молекул, встроенных в клеточную мембрану. Под воздействием даже небольшой деформации механочувствительный канал открывается и пропускает внутрь клетки поток положительно заряженных ионов. Ионный ток возбуждает нервное окончание, вернее, его мембрану, и, когда возбуждение достигает определенного порогового значения, на мембране возникает потенциал действия. Он распространяется по мембране аксона сенсорного нейрона и идет дальше клеточного тела к спинному мозгу, где сигнал передается через синапс второму нейрону и проводится им в головной мозг для обработки. Кожный сенсорный нейрон сообщает остальной нервной системе три вида информации: что-то прикасается к вашей коже; место прикосновения находится чуть выше правого запястья и это что-то довольно легкое.
Начнем с вопроса «Где?». Определить место воздействия раздражителя очень просто. Нервные окончания каждого тактильного нейрона охватывают конкретный ограниченный участок кожи, который может быть совсем крошечным, например, на руке или губе, или довольно большим, скажем, на спине. Поскольку мозг знает, где расположено рецептивное поле каждого нейрона, он мгновенно определяет, где именно происходит воздействие раздражителя[3]. Очевидно, что на кончике пальца, густо усеянном крошечными нервными окончаниями, локализация происходит гораздо точнее, чем на спине, где нервных окончаний намного меньше и каждый нейрон отвечает за гораздо бо́льший по площади участок кожи.
«Рецептивное поле» является важным понятием в науке о восприятии. На нашем рисунке рецептивное поле, охваченное терминальными веточками (нервными окончаниями) аксона сенсорного нейрона, представляет собой очерченный пунктирной линией овал. Здесь это участок кожи, воздействие на который приводит к возбуждению конкретного осязательного нейрона. Как вы увидите дальше, точно так же устроена и система зрительного восприятия: под рецептивным полем понимается участок сетчатки, возбуждающий конкретный зрительный нейрон на уровне самой сетчатки или на других уровнях зрительной системы.
Второй вопрос – «Насколько сильно?», то есть какова интенсивность стимула. Каким образом кожный сенсорный нейрон сообщает головному мозгу, что воздействие раздражителя является слабым или сильным? Все сенсорные нейроны – осязательные, слуховые, зрительные, обонятельные или вкусовые – кодируют эту информацию в виде частоты потенциалов действия. Легкое прикосновение генерирует всего несколько потенциалов действия, более сильное – быструю и частую последовательность таких импульсов. Благодаря этому наш мозг – или же исследователь, регистрирующий скорость активации нейронов, – может определить интенсивность воздействия раздражителя.
Многие ученые, включая меня, выдвинули предположение, что в конкретных паттернах потенциалов действия может быть закодирована дополнительная информация аналогично тому, как это делается в азбуке Морзе[4]. Этот паттерн может сообщать мозгу, например, о том, от какого типа рецептора данного аксона поступает сигнал (смотрите следующий параграф). Само собой разумеется, характер паттерна влияет на реакцию мозга: известно, что импульсы с короткими интервалами между ними возбуждают постсинаптические клетки намного сильнее, чем импульсы с более длинными интервалами. Тем не менее никто пока не предложил и не исследовал конкретную расшифровку этого кода.
Третий вопрос «Что?», пожалуй, самый интересный. Мозгу важно знать: «Что прикоснулось к моему запястью?» Не все прикосновения одинаковы, поэтому существует несколько разных типов осязательных нейронов, реагирующих на различные виды внешнего воздействия. Один тип тактильных рецепторов умеренно чувствителен к легким прикосновениям к поверхности кожи и посылает сигналы в мозг на протяжении всего времени, пока длится прикосновение. Другой тип рецепторов реагирует только на довольно сильное давление и только на его изменения – он посылает сигналы в мозг, когда давление начинается и когда заканчивается. На сегодняшний день нам известно более дюжины видов первичных осязательных нейронов. Каждый из них можно проверить на приеме у невролога – что, собственно говоря, он и делает, когда колет вас иголкой или прикасается к вам вибрирующим камертоном.
Интересно, что многие различия между кожными рецепторами обусловлены не фундаментальными различиями в их строении, а разными структурами, в которые встроены их нервные окончания. Нервные окончания осязательных нейронов окружены специализированными клеточными структурами, которые и определяют, на какие виды прикосновений реагирует осязательный нейрон. Представьте себе барабанные палочки для малого барабана и бас-барабана. Они отличаются между собой только тем, что у первых на ударном конце находится маленький деревянный шарик, а у вторых на этот шарик надет большой меховой наконечник. Из-за этого при ударе о натянутую кожу барабана они издают очень разные звуки. Кроме того, разнообразие реакций рецепторов обеспечивается и различиями в ионных каналах на их мембране. Эти детали, хотя и свидетельствуют об удивительном мастерстве эволюции, не особенно важны для нашего обсуждения. А важно то, что разные типы нейронов реагируют на разные внешние воздействия: одни способны почувствовать прыжок блохи, а другие реагируют лишь на удар кулаком. Разумеется, между этими крайностями существует множество промежуточных вариантов, и в большинстве случаев сигналы о воздействии стимула передаются в мозг смесью разных типов нейронов. Как замечательно выразился один из моих коллег, «подобно отдельным инструментам в оркестре, каждый подтип [осязательных нейронов] сигнализирует о той или иной конкретной характеристике действующего на кожу стимула, что в совокупности создает симфонию нервных импульсов, которые интерпретируются мозгом как осязательное ощущение»[5].
Это общий принцип функционирования всех сенсорных систем. Вкусовая система включает набор из пяти типов рецепторов, отвечающих за восприятие сладкого, кислого, соленого, горького вкусов, а также умами (сложного вкуса, присущего некоторым аминокислотам). Примечательно, что на сегодняшний день ученые насчитали в обонятельной системе около 400 типов рецепторов, каждый из которых обладает избирательной чувствительностью к конкретному пахучему веществу. Это объясняет способность дегустаторов различать сотни видов вин по их букету (к сожалению, я не обладаю этим талантом), а также то, почему аромат конкретных духов или одеколона способен вызвать воспоминания о бывшей любви.
ЧЕМ ЗРЕНИЕ ПОХОЖЕ НА ОСЯЗАНИЕ?
В предыдущем разделе я так подробно остановился на осязании, потому что основные принципы работы нашей осязательной и зрительной систем очень похожи. Их сенсорные нейроны, по сути, делают одно и то же. И зрение, и осязание сводятся к тому, чтобы передать в мозг информацию о стимулах, воздействующих на определенный участок пласта сенсорных клеток – в коже или сетчатке. Обе системы состоят из разнообразных типов рецепторов. В той и другой индивидуальные нейроны реагируют на стимулирование ограниченного рецептивного поля и сообщают мозгу конкретные виды информации. Но что касается зрения, то здесь мы знаем намного больше о том, как головной мозг принимает, обрабатывает и интерпретирует сигналы, поступающие от сетчатки глаза.
Как было сказано выше, отдельные иннервирующие кожу нейроны сообщают мозгу разные виды информации о воздействующих на них стимулах. Этот же фундаментальный принцип лежит в основе работы зрительной системы: каждое волокно зрительного нерва передает в мозг информацию об одном небольшом участке и одном конкретном аспекте видимого мира.
Сетчатка, по сути, представляет собой микропроцессор, подобный тому, что находится в вашем смартфоне, фотоаппарате или электронных часах. Она содержит множество разных типов нейронов, о которых дальше мы поговорим очень подробно. А пока давайте посмотрим, как сигналы с сетчатки передаются в головной мозг. Это делается посредством нейронов дальней связи, называемых ганглионарными клетками сетчатки (и аналогичных осязательным нейронам, идущим от кожи к спинному мозгу). Сетчатка человеческого глаза содержит около миллиона ганглионарных клеток. Они собирают входные сигналы от нескольких типов внутренних нейронов сетчатки и посылают их в мозг через зрительный нерв, который образован из соединенных в пучок длинных аксонов ганглионарных клеток.
Первое серьезное исследование ганглионарных клеток сетчатки было предпринято американским ученым венгерского происхождения Стивеном Куффлером. Хотя научный интерес Куффлера был сосредоточен на другом предмете – а именно на механизме синаптической передачи, судьба в какой-то момент свела его с офтальмологией. Немало попутешествовав по миру, после Второй мировой войны он получил место на кафедре офтальмологии Университета Джонса Хопкинса. Отчасти из благодарности к своим работодателям он провел экспериментальное исследование, которое по сей день остается фундаментальным для нашего понимания феномена зрения.
Примерно в 1950 г. Куффлер занялся изучением электрической активности одиночных ганглионарных клеток в сетчатке глаз кошек, находящихся под воздействием глубокого наркоза. Исследователи вводили в глаз животного микроэлектрод, подводили его к ганглионарной клетке, после чего регистрировали генерируемые клеткой последовательности импульсов при стимуляции поверхности сетчатки пятнами света. Пятна света должны были быть очень мелкими, чтобы имитировать изображения объектов внешнего мира, которые, как известно, отображаются на сетчатке в сильно уменьшенном виде – например, изображение моего ногтя большого пальца на расстоянии вытянутой руки на сетчатке не превышает в размере 0,4 мм.
Куффлер заметил, что сигналы ганглионарных клеток сетчатки очень похожи на сигналы сенсорных нейронов кожи. Каждая ганглионарная клетка отвечала за один небольшой участок поверхности сетчатки – свое рецептивное поле. Самая маленькая рецептивная зона в кошачьем глазу имела размер около 40 мкм или 0,04 мм. Хотя никто никогда не измерял (по медицинским соображениям) размер отдельных рецептивных полей у людей, косвенные свидетельства указывают на то, что их минимальный диаметр составляет около 10 мкм. Как рассчитал один лауреат Нобелевской премии, 10-микрометровое рецептивное поле соответствует изображению 25-центовой монеты, полученному с расстояния около 150 м. Лично я не способен увидеть четвертак, находящийся так далеко, но, возможно, у нобелевских лауреатов более острое зрение, чем у простых смертных. Рецептивные поля можно сравнить с пикселями на компьютерном мониторе. Чем больше плотность ганглионарных клеток, тем острее зрение.
НЕМНОГО КОНТЕКСТА
На заре нейробиологии – примерно с 1945 по 1980 г. – самые захватывающие исследования проводились в области регистрации электрических сигналов. Эти сигналы включали мозговые волны, которые записывались с помощью наложенных на поверхность головы электродов (в виде электроэнцефалограмм или ЭЭГ) и дистанционно отражали общую электрическую активность мозга, а также сигналы, которые фиксировались посредством введения в мозг тонких микроэлектродов и свидетельствовали об активности отдельных нейронов. Изучение электрической активности мозга было, как говорится, самой крупной игрой в городе. (Молекулярная генетика, которая сегодня стала главным двигателем биологической науки, тогда была представлена в основном биохимией, а генная инженерия еще не появилась.)
Вряд ли стоит говорить, что электрические сигналы отдельных нейронов чрезвычайно слабы, а потому уязвимы для помех со стороны всех видов проходящих мимо электромагнитных волн, излучаемых полицейскими рациями, телевизионными вышками, медицинскими пейджерами и т. д. Поэтому испытуемого, человека или животное, часто помещали в специальную проволочную «клетку», которая экранировала нежелательные электромагнитные волны.
Другим способом защититься от помех было поместить между их источником и местом записи непроницаемую преграду – например, землю. Вот почему многие лаборатории размещались в подвалах или же их стены обшивались специальными медными экранами. (С тех пор регистрирующая аппаратура значительно улучшилась, усовершенствованные усилители позволяют нам чисто записывать самые слабые сигналы, поэтому необходимость в таких крайних мерах защиты сегодня отпала.)
В те времена в типичной исследовательской лаборатории работали три-четыре исследовательские группы, каждая из которых занимала три-четыре комнаты. Группа включала независимого руководителя (профессора), а также трех-четырех постдоков (молодых ученых, недавно получивших докторскую степень) и лаборантов. Профессора обычно ютились в крошечных угловых кабинетах, постдоки – в отдельной комнате или закутке рядом с аппаратной. Виварий для мелких млекопитающих обычно располагался в конце длинного коридора. В первый день резкая вонь буквально валила новичка с ног. К счастью, запах постепенно ослабевал и через несколько недель пропадал вовсе. Мыши и кролики с их подстилками и экскрементами никуда не исчезали, но ваша обонятельная система через какое-то время просто переставала реагировать на этот раздражитель (о, благословенный дар сенсорного привыкания!).
В этих лабораториях почти не было стеклянных колб и бутылей, которые так любят показывать в фильмах. Вместо них всюду стояли стеллажи с усилителями, акустическими колонками, записывающими устройствами, блоками питания и прочей электроникой. Если лаборатории посчастливилось иметь компьютер, тот был размером с холодильник, имел мощность меньше моего смартфона и понимал только машинный язык. Программировать его умел лишь специально обученный программист, который использовал для этого примитивный машинный код, недалеко ушедший от двоичных последовательностей нулей и единиц. Жужжащие вентиляторы, охлаждавшие легко перегревавшиеся внутренности электронного монстра, смешивали запах новой проводки и горячего металла с запахами животных, спирта и эфира.
Мы невероятно дорожили лабораторным оборудованием. Нашей главной рабочей лошадкой был катодно-лучевой осциллограф с тускло светящимся зеленым экраном, дедушкой современных компьютерных дисплеев. Снимки экрана мы делали на пленочный фотоаппарат. Осциллограф требовал тщательной калибровки, и, поскольку он, как старый радиоприемник, работал на электронных лампах, с утра первым делом мне нужно было не забыть его включить, чтобы он успел нагреться к тому времени, когда мы планировали начать работу. Когда я создал собственную лабораторию, первый осциллограф обошелся нам в 2500 баксов (в долларах 1970-х гг.). Сегодня хороший аппарат можно купить всего за 500 долларов.
СТИВЕН КУФФЛЕР
Основоположник нейробиологии Стивен Куффлер сделал очень много для того, чтобы эта наука стала такой, какой мы ее знаем сегодня. Огромную роль сыграли не только его новаторские научные работы и публикации, приверженность делу и высокая планка, которую он задавал для всех окружающих, но и умение выбирать людей – учеников и коллег, которые сегодня составляют значительную часть элиты в американской нейрологии. Его боготворили все, кто его знал. Техники, обслуживающие оборудование, секретари, лучшие ученые умы – все обожали Стивена Куффлера[6].
Худощавый и подвижный, он напоминал подростка. Всю жизнь он увлекался теннисом, но мало кто знал, что в юности он был чемпионом Австрии в этом виде спорта. Куффлер родился в 1913 г. в Венгрии и вырос в семейном имении. В автобиографии он назвал это место фермой, но другие описывали его как крупное поместье, где работало большинство жителей соседней деревни. Раннее детство было счастливым временем, но в 1919 г., когда в Венгрии разразилось коммунистическое восстание, его семье пришлось бежать в Австрию. Куффлер учился в католической школе-интернате, а затем поступил в медицинскую школу. К несчастью, его отец разорился и вскоре умер, оставив семью без средств. Юному Стивену, которому на тот момент не исполнилось еще и 20 лет, пришлось самому зарабатывать себе на жизнь. В 1937-м он окончил медицинскую школу и снова бежал из страны, едва успев до германского вторжения в Австрию.
Через Венгрию, а затем через итальянский Триест Куффлер перебрался в Лондон, где у него были друзья. Но, не имея лицензии на медицинскую практику в Англии, он был вынужден снова эмигрировать, на этот раз в Австралию, где познакомился с Джоном Экклсом и Бернардом Кацем, будущими титанами нейробиологии, и начал свою карьеру ученого-исследователя. В период интенсивной работы с 1939 по 1944 г. эти трое сделали фундаментальные открытия в области нейронной проводимости и работы синапсов.
Но административные препоны положили конец сотрудничеству. Куффлер со своей австралийской невестой уехал в Чикаго. Его научная репутация уже бежала впереди него. В конце концов он перебрался в Гарвард, где основал кафедру нейробиологии – если это была и не первая академическая кафедра нейронаук в мире, то наверняка одна из первых. На тот момент такой научной дисциплины еще не существовало, а Американское общество нейронаук было создано лишь пару десятилетий спустя (мой членский номер 000064 выдает во мне настоящего старожила). Кафедра Куффлера быстро стала ведущей в Северной Америке. Она славилась семейной атмосферой, а студентов туда тщательно отбирали. Через несколько лет после ее основания я провел там два года как приглашенный научный сотрудник.
Исследовательские стандарты на кафедре чрезвычайно высоки: там царил дух научной элитарности, если не сказать высокомерия, и они не стыдились этого. В некотором роде кафедра Куффлера была академическим эквивалентом «Волшебного автобуса» Кена Кизи[7]. Если вы соответствовали их критериям, вас принимали в семью. Если нет, так прямо и говорили.
Атмосфера в этом научном «автобусе» была потрясающей; ученые развлекались от души. Сам Куффлер любил пошутить и прославился своей простотой и жизнерадостностью во всем Гарварде. Сотрудники его кафедры были Веселыми шутниками нейронауки, представляя собой странную и уникальную смесь высочайшего интеллекта, жесткой дисциплинированности, требовательности и умения подурачиться. Семинары проходили весело; от докладчиков ожидали, что их выступления будут безукоризненными с научной точки зрения, но при этом доходчивыми и приправленными здоровой долей юмора.
На кафедре имелась своя обеденная комната, где ученые обычно отдыхали в перерывах между экспериментами. Так со временем родилась практика обеденных семинаров. По мере того как известность кафедры росла, в Бостон приезжало все больше любопытствующих ученых, и их обычно приглашали провести семинары. В конце концов гостей оказалось так много, что запланировать отдельное мероприятие для каждого стало попросту невозможным. Решением стала система обеденных семинаров. Все, что требовалось сделать приглашающему сотруднику, – написать имя докладчика на календаре на двери комнаты. Не было никакого формального отбора и утверждения докладчиков, никаких официальных программ; вся ответственность лежала на приглашающем. Тем не менее контроль качества существовал, и довольно жесткий: если ты приглашал плохого докладчика, ты терял лицо. Хуже того, твоего гостя могли публично разнести в пух и прах.
На этих обеденных семинарах на нас обрушивалась лавина захватывающих научных новостей. Два-три таких выступления в неделю позволяли нам полностью быть в курсе того, что происходило за стенами нашей лаборатории. О многих новых открытиях мы зачастую узнавали еще до их публикации, что на практике давало нам весомое конкурентное преимущество. Мы шли в авангарде нейронауки и невероятно гордились этим.
Физически доступ посторонним на кафедру не был запрещен (в университетах это не поощряется), но фактически ее мир был закрыт для чужаков. Никто не захлопывал перед ними дверь, но и никогда не приглашали на внутренние мероприятия, тем более на обеденные семинары. Если кому-то хватало глупости явиться по собственной инициативе, его ожидал ледяной прием. Естественно, «люди со стороны» – другие гарвардские ученые – возмущались своим положением аутсайдеров, особенно когда видели через стеклянную стену, как сотрудники кафедры нейробиологии смеются, что-то увлеченно обсуждают и занимаются своей удивительной наукой. Но для группы Куффлера это было волшебное время. Так продолжалось до его смерти в 1980 г., после чего коллектив на удивление быстро распался. Сегодня кафедра нейробиологии Гарвардского университета по-прежнему остается замечательным местом и мировым лидером в области нейронаук, но ее бывшие сотрудники с ностальгией вспоминают славные былые времена.
(Если честно, все было не так однозначно. Необходимость показать себя с лучшей стороны слишком давила на сотрудников. Как признался мне один из ветеранов кафедры Куффлера, это был потрясающий опыт и ему понадобилось всего два года психотерапии, чтобы оправиться от него. Кроме того, нередкий в науке авторитарный стиль руководства иногда приводил к ошибкам.)
Как Стив Куффлер, этот обаятельный человек, брызжущий плохими каламбурами, сумел оказать такое влияние? После его смерти друзья и ученики опубликовали сборник воспоминаний. В нем Гюнтер Стент, отец-основатель молекулярной биологии, назвал Стива неподкупным. Другие превозносили научные таланты и принципиальность Куффлера, но характеристика «неподкупный» говорит о многом – о душевной чистоте.
Куффлер отвергал любую помпезность, предпочитая ей приземленность. Как-то он засиделся допоздна за пинтой пива в соседнем пабе в компании пары постдоков, доцента и Торстена Визеля, будущего лауреата Нобелевской премии, сменившего его на посту заведующего кафедрой. Когда Визель принялся ворчать насчет бремени административной рутины, Стив посмотрел на него и с легкой улыбкой заметил: «Если хочешь прославиться, тебе придется пахать как проклятому».
Однажды за ланчем Стив услышал, как я сетую коллегам о том, что научная тема, над которой я работаю, была слишком узкой, а не общей (в те времена мне казалось, что она имеет отношение к сетчатке и только). Стив сидел на другом конце стола и спокойно ел свой обед из пластикового контейнера. Посмотрев на меня, он улыбнулся и просто сказал: «Мы изучаем общее через изучение конкретного».
ЦЕНТРАЛЬНОЕ И ПЕРИФЕРИЧЕСКОЕ ЗРЕНИЕ
Чтобы понять, как сетчатка формирует визуальное сообщение для головного мозга, необходимо сначала узнать, как организованы ее нейроны. Сетчатка не просто набор светочувствительных клеток. Она содержит пять основных классов нейронов, каждый из которых выполняет свою функцию. Первый класс – фоторецепторы, называемые палочками и колбочками (мы говорим, что с них начинается зрительный анализатор).

Эти фоторецепторные нейроны чувствительны к свету (благодаря колбочкам мы видим в период от рассвета до заката, благодаря палочкам – при свете луны и звезд) и являются основными функциональными клетками сетчатки. Палочки и колбочки соединяются через синапсы с особым типом интернейронов – биполярными клетками, называемыми так потому, что, в отличие от других нейронов, у них имеются два четких полюса: один для получения, другой для передачи сигналов. Биполярные клетки получают входные сигналы от палочек и колбочек и передают их на ганглионарные клетки, чьи длинные аксоны, соединяясь в пучок, образуют зрительный нерв. Именно через ганглионарные клетки сетчатки мозг получает всю информацию о визуальном мире.
Чуть позже мы рассмотрим еще два типа нейронов сетчатки, которые делают процесс зрительного восприятия еще более захватывающим. Но пока вам необходимо понять, что фоторецепторы, биполярные и ганглионарные клетки составляют основу сетчатки и их организация определяет остроту нашего зрения.
Чтобы увидеть 25-центовую монету на расстоянии 150 м, требуется очень острое зрение. За него отвечает особая область сетчатки, называемая центральной ямкой, или фовеа. Большинство людей знают, что их периферическое зрение хуже центрального, но редко осознают, насколько велика эта разница. У обычного человека острое центральное зрение охватывает сектор всего около пяти градусов, что эквивалентно половине ширины моей ладони на расстоянии вытянутой руки. За его пределами острота зрения резко снижается. На самом деле, если я отведу свою вытянутую руку в сторону на 30–60 см, я с трудом смогу сосчитать на ней пальцы. У врачей-офтальмологов в обиходе имеется даже специальный термин для характеристики плохого зрения – «острота зрения, равная счету пальцев» (finger-counting acuity). Следующая ступень плохого зрения – когда пациент видит «только движения руки». Другими словами, в центральном поле мы видим хорошо, а за его пределами – почти слепы.
Почему же мы практически не осознаем, насколько слабо наше периферийное зрение? Когда наши глаза сканируют пространство, нам кажется, что мы видим объекты гораздо четче, чем они (как показывают измерения) видятся нам на самом деле. Это может быть связано с тем, что в нашей зрительной памяти зафиксированы все объекты, которые мы однажды детально рассмотрели центральным зрением.
Но наше периферийное зрение далеко не бесполезно. Оно выполняет как минимум две важные функции. Во-первых, оно очень чувствительно к изменениям на периферии поля обзора. То, что внезапно появляется, движется или мигает, мгновенно привлекает наше внимание к этому месту, побуждая исследовать его центральным зрением.
Во-вторых, периферийное зрение помогает нам ориентироваться в пространстве. Когда мы движемся, периферийным зрением мы отслеживаем крупные объекты: дверной проем, диван, холодильник, другого человека. Хотя эти изображения не детализированы, это позволяет нам прокладывать путь между препятствиями, избегая столкновения с ними. Данная функция периферийного зрения особенно наглядно проявляется у пациентов с возрастной макулярной дегенерацией (заболеванием сетчатки, которым страдает около 15 % белых американцев в возрасте 80 лет и старше). Из-за частичного разрушения нейронов в центральной ямке у таких больных значительно ухудшается центральное зрение, вплоть до полной его потери, однако периферийное зрение почти сохраняется. В результате они не могут читать, узнавать лица или смотреть телевизор, но по-прежнему в состоянии передвигаться в пространстве – по своему дому и даже по городским улицам. Хотя с медицинской точки зрения они считаются слепыми, посторонний наблюдатель может не сразу это заметить.

Эта разница между центральным и периферийным зрением объясняется очень просто: в центральной области нашей сетчатки плотность пикселей гораздо выше, чем на периферии (см. рисунок). Пиксели, которые имеют значение в данном случае, – это ганглионарные клетки, последние звенья в нейронной цепи сетчатки, аксоны которых образуют зрительный нерв, идущий в головной мозг. На рисунке выше отдельные ганглионарные клетки обозначены черными кружками, а Т-образные структуры над ними показывают рецептивные поля, с которых они собирают входные зрительные сигналы (другие клетки сетчатки на рисунке не показаны). В центральной части сетчатки самая высокая численность – и, следовательно, плотность расположения – ганглионарных клеток, вследствие чего каждая клетка имеет очень малое рецептивное поле. В направлении от центра к периферии ганглионарные клетки расположены все реже, а их рецептивные поля становятся все больше. Чем больше участок сбора входных зрительных сигналов, тем крупнее пиксели и тем ниже качество изображения.
Почему эволюция наделила значительную часть поверхности нашего глаза таким слабым зрением? Почему бы не покрыть всю сетчатку плотным слоем ганглионарных клеток, чтобы мы могли хорошо видеть не только ее центральной, но и периферической частью? Главное достоинство устройства нашей сетчатки опять же в экономичности. Ганглионарные клетки сетчатки – дорогое удовольствие. Они занимают место не только в сетчатке, но и в зрительном нерве: каждая такая клетка должна протянуть в мозг свой аксон. В норме зрительный нерв человека имеет диаметр около 4 мм. Но если бы плотность ганглионарных клеток по всей сетчатке была такой же, как в центре, то наш зрительный нерв был бы толщиной с садовый шланг. Не говоря уже о прочих моментах, мы бы попросту не могли двигать глазом в глазнице.
Кроме того, посылать в мозг такой плотный поток информации имело бы смысл только при условии, что мозг был бы в состоянии ее обработать и найти ей полезное применение. Как бы выглядел мир, если бы вы видели все поле обзора так же четко, как пространство вокруг точки фиксации? С одной стороны, было бы потрясающе видеть мир таким, каким он предстает на высококачественной фотографии (бо́льшую часть которой вы можете охватить центральным зрением). Но что бы вы делали со всей этой информацией? Могли бы вы ее обработать всю сразу?
Похожая стратегия используется в некоторых управляемых бомбах и других военных устройствах с оптическим наведением. Их создатели не любят разглашать детали, но известно, что для локализации интересующего региона используется низкое разрешение, после чего разрешение увеличивается, чтобы максимально детализировать изображение и обнаружить искомую цель – используя минимально возможное количество вычислительного оборудования и ресурсов.
ПОЧЕМУ У ЯСТРЕБА ЯСТРЕБИНОЕ ЗРЕНИЕ?
Теперь давайте совершим путешествие в царство животных. Представьте себе пшеничное поле в конце лета после уборки урожая. Оно покрыто стерней и остатками соломы коричневато-песочного цвета. У самой земли копошатся полевые мыши, кормясь пшеничными зернами, выпавшими из колосьев во время жатвы. В небе на высоте около 60 м грациозно парит ястреб. Вдруг он складывает крылья и пикирует вниз. Когда он снова взмывает в небо, в когтях у него зажата мышь.
Как ястреб сумел с такой высоты разглядеть крохотную мышь в пять сантиметров длиной, прячущуюся среди стерни? У мыши неприметная коричневато-песочная шкурка под цвет соломы, а ястреб летит с довольно большой скоростью. Недаром люди называют очень острое зрение ястребиным. Многие пытались объяснить, в чем причина такой способности, выдвигая разные гипотезы. Например, что колбочковые клетки – светочувствительные рецепторы – у ястребов тоньше, чем у большинства других животных, поэтому они расположены в сетчатке более плотно. Или что у ястребов большое поле зрения – 290° по сравнению с 180° у людей. У ястребов большие глаза: они занимают гораздо бо́льшую часть его маленькой головы, чем глаза у человека или какого-либо другого млекопитающего. Большие глаза – это хорошо; как известно, чем больше объектив фотоаппарата, тем выше четкость фотографий. (Профессиональные фотографы, снимающие футбольный матч на краю поля, используют такие огромные объективы, что иногда с трудом удерживают фотоаппараты в руках, а некоторые камеры им приходится ставить на треногу.)
Все это интересные гипотезы, но в большинстве случаев в них присутствует элемент предвзятости. Авторы этих исследований знают, что ястребы хорошо видят, поэтому просто пытаются объяснить этот факт, вместо того чтобы объективно исследовать ястребиный глаз. Некоторые из объяснений не выдерживают проверки. Например, колбочковые клетки у ястребов действительно маленькие и размещены с большой плотностью, но их плотность всего примерно на 60 % выше, чем у человека. Кроме того, глаз у ястреба действительно большой для такого маленького животного, составляя в диаметре около 12 мм, но в два раза меньше человеческого (24 мм). Конечно, у людей голова намного больше, но это никак не влияет на физику света: что касается оптики, то здесь люди далеко впереди.
Наконец, сравним разрешающую способность человеческого и ястребиного глаза[8]. При должном терпении можно научить ястреба выбирать между объектом-мишенью с лакомством (например, раскрасив мишень узкими полосами) и пустой мишенью (с широкими полосами), после чего протестировать, насколько тонкие полосы он способен различать. У пустельги, лучше всего изученного вида ястребов[9], разрешение зрения на самом деле несколько хуже, чем у людей.
Но постойте! Как же тогда ястреб ловит неприметных пятисантиметровых мышей среди стерни на сжатом поле? Нет никаких сомнений в том, что он видит гораздо лучше, чем мы. Как разрешить это кажущееся противоречие?
Я не ставлю под сомнение наблюдения орнитологов. Но на мой взгляд, главная причина превосходного зрения ястреба в том, что он хорошо видит всей сетчаткой, а не только ее центральной частью. Доказательством тому может служить количество и распределение нейронов сетчатки. Колбочковые клетки у ястреба не могут быть расположены намного плотнее, чем у нас: есть предел тому, сколько рецепторов можно втиснуть в ограниченное пространство. Но что действительно определяет остроту зрения, так это плотность ганглионарных клеток, а не колбочек.
Ключевой принцип состоит в том, что разрешение любой системы передачи информации ограничено плотностью размещения наименее густо расположенных элементов в системе. В сетчатке глаза – человека или ястреба – такими элементами являются ганглионарные клетки, которые составляют всего несколько процентов от общего количества нейронов в любом глазу. Известно, что у большинства животных их плотность резко снижается на периферии сетчатки. У ястребов такой перепад гораздо слабее выражен. На самом деле количество ганглионарных клеток в сетчатке ястребов в несколько раз превышает таковое у людей – около восьми миллионов по сравнению примерно с одним миллионом у среднестатистического человека. И эти ганглионарные клетки занимают у ястреба меньшую площадь. Да, у ястребов толстый зрительный нерв, но это им не мешает, потому что они больше используют движения головы, чем движения глаз.
Каким образом задействуются все эти ганглионарные клетки? Прежде всего в ястребином глазу, как у многих других птиц, имеется две области острого зрения – центральные ямки, одна из которых находится в центре, а другая – чуть сбоку (у нас всего одна). Но главное – это общее распределение ганглионарных клеток по сетчатке. У человека их плотность на дальней периферии составляет всего 1 % от их плотности в центре. У ястребов эта разница гораздо меньше – например, у пустельг периферическая плотность ганглионарных клеток составляет 75 % от центральной, то есть на периферии сетчатки у них насчитывается около 15000 ганглионарных клеток на квадратный миллиметр, тогда как у людей – всего 500. Если мы практически слепы на периферии, то ястребы – нет. Мыши трудно скрыться от их острого взора, способного сканировать полосу поля шириной в несколько метров.
Как я заметил несколько абзацев назад, неизвестно, как бы человеческому мозгу удавалось обрабатывать колоссальный поток зрительной информации, будь наше зрение одинаково острым по всей сетчатке. Похоже, ястребы находятся именно в такой ситуации – так как же они с этим справляются? Мы можем только догадываться, но ответ, скорее всего, кроется в том, что в их мозгу имеется мощнейший компьютер для обработки визуальных данных. Значительную часть ястребиного мозга занимает так называемое верхнее двухолмие – мозговая структура, которая также есть у людей. Пока мы не знаем, как именно работает эта нейронная система, но исходя из ее размера можем предположить, что она существенно превосходит в своей производительности подкорковые зрительные структуры человеческого мозга. Когда-нибудь, когда секреты обработки визуальной информации в зрительной системе птиц будут раскрыты, эти принципы могут быть применены и к искусственным системам обработки изображений. Создатели Adobe Photoshop, вам есть чему поучиться у пернатых!
3 | Микропроцессор в глазу
Человек склонился над гитарой,Непохожий на других, почти изгой.День был зелен,И ему сказали: «Голубой своей гитаройИскажаешь мир ты – он совсем другой».А в ответ он: «Но моя гитара голубаяИзменяет мир тот, что мы знаем».УОЛЛЕС СТИВЕНС
Итак, мы выяснили, что лучше всего видим частью глаза с высокой плотностью нейронов. Но не все ганглионарные клетки сетчатки одинаковы. Это не стандартные фотоэлементы вроде тех, что используются в приборах («магических глазах»), которые обнаруживают проникших в ваш дом грабителей или не дают закрыться дверям лифта, пока вы не вошли в него или не вышли. Ганглионарные клетки по-разному реагируют на разные визуальные раздражители – аналогично тому, как это делают разные типы осязательных нейронов в коже. Видимая картина фрагментируется, раскладывается на наборы специализированных сигналов. Этот начальный этап обработки изображения влияет на то, как мы видим восход солнца, как уворачиваемся от выскочившего из-за угла автомобиля, как узнаем лицо любимого человека и как наслаждаемся полотнами Ван Гога.
ОБРАБОТКА ИЗОБРАЖЕНИЙ – 1: СЕТЧАТКА РАСКЛАДЫВАЕТ ИЗОБРАЖЕНИЕ НА СОСТАВЛЯЮЩИЕ
Начнем мы с самого простого типа кодирования, основанного на различиях между ганглионарными клетками с устойчивым и транзиторным ответами. Некоторые ганглионарные клетки сетчатки, реагируя главным образом только на появление стимула, генерируют короткую серию импульсов (пачку спайков) – это клетки с транзиторным ответом. Клетки с устойчивым ответом посылают сигналы в мозг на протяжении всего времени действия стимула. Как вы помните, точно таких же два способа сигнализации используются осязательными нейронами, чтобы сообщить мозгу о прикосновениях к вашей коже.
На разницу между устойчивым и транзиторным ответом налагается еще одно важное различие: одни ганглионарные клетки под действием светового стимула возбуждаются, генерируя короткую или устойчивую череду импульсов, – это клетки с так называемым on-ответом (от английского on – «включено»); другие тормозятся – это клетки с off-ответом (от английского off – «выключено»). Таким образом, мы получаем четыре типа ганглионарных клеток:
● клетки с транзиторным on-ответом;
● клетки с транзиторным off-ответом;
● клетки с устойчивым on-ответом;
● клетки с устойчивым off-ответом.
Что это значит для нашего зрения? Представьте, что вы головной мозг. Ваша задача – на основе последовательности потенциалов действия, поступающих через зрительный нерв, определить, какое событие произошло во внешнем мире.
Клетка с транзиторным ответом реагирует главным образом на первое появление визуального стимула, после чего почти замолкает. По сути, это детектор изменений. Сигналы этого типа клеток почти бесполезны для распознавания лиц в толпе: клетки активизируются всего на несколько сотен миллисекунд – мгновение, за которое глаз не успевает зафиксировать форму глаз, носа, рта и т. д. Поэтому в данном случае ваш мозг полагается на клетки с устойчивым ответом, которые передают ему непрерывный поток информации, когда вы задерживаете взгляд на лице. Теперь представьте другую ситуацию: где-то в вашем поле зрения вдруг мелькает птеродактиль. Ваша сетчатка должна максимально быстро и мощно сообщить об этом мозгу. Это обязанность клеток с транзиторным ответом. Они молчат бо́льшую часть времени, но мгновенно вспыхивают и выстреливают интенсивной пачкой импульсов, чтобы предупредить мозг о внезапном появлении объекта в своем рецептивном поле. Рекламщики знают, что мигающее изображение действует намного сильнее, чем неподвижное, и клетки с транзиторным ответом объясняют почему.
Кроме того, одни клетки реагируют на освещение, другие – на затемнение. С освещением все понятно, но с реагированием на затемнение немного сложнее. Эти два типа реакции называются on-ответом и off-ответом соответственно.
Как было сказано выше, одни ганглионарные клетки активизируются в ответ на увеличение освещенности в пределах их рецептивного поля – их называют on-клетками. Другие – off-клетки – реагируют на ослабление освещенности. Как функционирует данный механизм? Дело в том, что практически каждый видимый объект состоит из более светлых и более темных частей. Рассмотрим простой пример – полосу из двух частей, белой и черной:  Представьте, что вы увлеченно читаете этот текст, как вдруг ваш взгляд останавливается на границе между белой и черной полосами. Какие сигналы ганглионарные клетки сетчатки посылают в ваш мозг? В то же мгновение, когда ваш взгляд фиксируется на этой границе, группа ганглионарных клеток с транзиторным on-ответом, отвечающая за восприятие области слева от точки фиксации, посылает в мозг интенсивный поток – пачку – потенциалов действия. Они сообщают вашему мозгу, что в их рецептивном поле появился более светлый, чем фоновая освещенность, объект. Одновременно с этим другая группа ганглионарных клеток – с транзиторным off-ответом, отвечающая за восприятие области справа от точки фиксации, – так же внезапно замолкает.
Представьте, что вы увлеченно читаете этот текст, как вдруг ваш взгляд останавливается на границе между белой и черной полосами. Какие сигналы ганглионарные клетки сетчатки посылают в ваш мозг? В то же мгновение, когда ваш взгляд фиксируется на этой границе, группа ганглионарных клеток с транзиторным on-ответом, отвечающая за восприятие области слева от точки фиксации, посылает в мозг интенсивный поток – пачку – потенциалов действия. Они сообщают вашему мозгу, что в их рецептивном поле появился более светлый, чем фоновая освещенность, объект. Одновременно с этим другая группа ганглионарных клеток – с транзиторным off-ответом, отвечающая за восприятие области справа от точки фиксации, – так же внезапно замолкает.
Да, именно так: мозг получает два потока сигналов, передающих одно и то же сообщение. On-клетки предупреждают мозг о появлении чего-то яркого слева от точки фиксации посредством всплеска импульсов; off-клетки передают то же сообщение – «Здесь появился светлый объект» – посредством снижения электрической активности.
Через несколько десятков миллисекунд ситуация меняется. Клетки с транзиторным ответом сделали свое дело и практически замолкают. Теперь в игру вступают клетки с устойчивым on-ответом: они генерируют стабильный поток потенциалов действия до тех пор, пока ваш взгляд фиксируется на границе между черным и белым. Клетки с устойчивым off-ответом на протяжении всего этого времени тормозятся. Роль ганглионарных клеток с устойчивым ответом очень важна: если бы в нашей сетчатке присутствовали только клетки с транзиторным ответом, светлый объект становился бы для нас невидимым через несколько десятков миллисекунд после попадания в наше поле зрения. Именно клетки с устойчивым ответом дают нам возможность рассмотреть мельчайшие детали внешнего мира, восприятие которых требует чуть больше времени, – и таким образом наделяют нас тем, что мы называем острым зрением.
Одновременно ганглионарные клетки, отвечающие за восприятие области справа от границы, посылают в мозг противоположные сигналы. Ваши клетки с транзиторным off-ответом сигнализируют в мозг, что в пределах их рецептивного поля появился более темный, чем фоновая освещенность, объект; клетки с транзиторным on-ответом реагируют противоположным образом. Через долю секунды эти сигналы пропадают, но клетки с устойчивым ответом продолжают передавать в мозг сообщение: «Этот темный объект все еще там» (on-клетки делают это своим способом, off-клетки – своим, посредством снижения электрической активности). Таким образом, сетчатка посылает в мозг мощный поток сигналов, когда светлый либо темный объект попадает в наше поле зрения: on-клетки реагируют на вкусную рыбку, мерцающую чешуей в темной воде, а off-клетки – на тень птеродактиля с когтистыми лапами, тихо скользящего на вас сверху.
ОБРАБОТКА ИЗОБРАЖЕНИЙ – 2: СЕТЧАТКА УЛУЧШАЕТ ИЗОБРАЖЕНИЕ РЕАЛЬНОГО МИРА
Клетки сетчатки выполняют еще одну важную функцию: они делают четче границы (контуры) объектов в передаваемом в мозг изображении. Обратите внимание, что on-клетки и off-клетки не трансформируют визуальное изображение; они просто сообщают мозгу о светлых и темных его частях. Улучшение границ – совсем другое дело, поскольку исходное изображение передается в мозг не совсем таким, какое оно есть на самом деле. С точки зрения мозга это существенное улучшение, поскольку границы объектов – место, где происходит основное действие и имеется максимум полезной информации.
То, что границы объектов важны, кажется очевидным. Но мало кто знает, что они воплощают в себе фундаментальный принцип, определяющий очень многие аспекты зрительного восприятия. Пиксели в зрительной картине окружающего мира расположены не случайным образом. Внешний мир проявляется в структурах – линиях, углах, кривых, поверхностях. Это означает, что визуальное содержание отдельных пикселей в значительной степени определяется их окружением. По-настоящему случайный визуальный мир выглядел бы как снежащий телеэкран. Наша зрительная система устроена таким образом, чтобы выделять структуры, в которых что-то меняется, и меньше фокусироваться на пространствах, где мало что происходит, – например, на внутренней части однотонной поверхности или на небе.
Такое улучшение границ осуществляется благодаря механизму, известному как латеральное торможение[10]. Это фундаментальный процесс для сетчатки глаза, а также для компьютерного зрения. Вернемся еще раз к примеру с черно-белой полосой. Срединные области сплошного черного или белого цвета не содержат много информации. Наибольшее количество информации несет граница между черной и белой областями. Так вот, механизм латерального торможения увеличивает интенсивность реакции ганглионарных клеток, чьи рецептивные поля находятся рядом с этой границей. Благодаря его действию разница между сигналами, поступающими в мозг от «приграничных» областей черной и белой зон, становится гораздо более выраженной, чем могла бы быть в ином случае. Это наглядный пример того, как сетчатка выбирает наиболее важные характеристики визуального мира, о которых следует сообщить мозгу.
В графических редакторах на наших компьютерах и смартфонах для корректировки цифровых изображений используются похожие функции, такие как «повысить контрастность» или «выделить контуры». Применение этих инструментов делает изображение более четким, хотя и, разумеется, не без издержек: за это приходится платить в той или иной мере потерей полутонов. Но иногда оно того стоит.
* * *
Механизм латерального торможения используется во всех сенсорных системах: не только в зрительной, но и в осязательной, слуховой, а также, как предполагают, в обонятельной и вкусовой. Он обнаружен у всех млекопитающих и многих видов беспозвоночных. В этом нет ничего удивительного. Широкое распространение того или иного полезного признака характерно для ранних стадий эволюции: латеральное торможение было одним из первых изобретенных природой эффективных приемов обработки сенсорной информации. Но почему увеличение контрастности контуров так полезно для живых организмов?
Чтобы ответить на этот вопрос, давайте рассмотрим, как механизм латерального торможения влияет на сигналы, посылаемые в мозг всей популяцией ганглионарных клеток сетчатки. На приведенном ниже рисунке показано, как фактическое изображение, падающее на поверхность сетчатки (и воспринимаемое палочками и колбочками фоторецепторов), преобразуется в модифицированный ответ, который передается в мозг ганглионарными клетками.

Вверху показано фактическое изображение: одна его половина – черная, а другая – белая. Внизу приведена амплитуда сигнала, посылаемого ганглионарными клетками в мозг. Обратите внимание: рядом с границей между черной и белой полосами сила сигнала меняется: на белой стороне она возрастает, а на черной, наоборот, снижается еще больше. Для мозга значит, что разница между светлой и темной областями – перепад в сигналах, определяющий наличие границы, – становится более выраженной.
Для простоты в вышеприведенном примере я говорил так, как если бы сетчатка содержала только on-клетки, тогда как в действительности примерно половину всех ганглионарных клеток составляют off-клетки. Их поведение противоположно поведению on-клеток, но общий эффект тот же – увеличение силы дифференциального сигнала рядом с границей. Я не буду утомлять вас пошаговым описанием процесса – у off-клеток все происходит так же, как у on-клеток, только наоборот.
Просто ради интереса задумайтесь вот над чем: если темная область стимула абсолютно черная, а белая область абсолютно белая, значит ли это, что механизм латерального торможения приводит к тому, что черная область рядом с границей выглядит чернее черной, а белая область – белее белой? Теоретически, если темная область стимула идеально черная, а белая – идеально белая, on– и off-клетки по определению функционируют на пределе своих возможностей – они не могут выйти за пределы нуля или 100 %. Но в реальном мире такого не бывает; все части изображения по степени освещенности или затемненности обычно находятся где-то посредине между абсолютными крайними точками. Когда наша зрительная система обнаруживает переход между более светлой и более темной областями, латеральное торможение усиливает дифференциальный сигнал и тем самым – наше восприятие контраста. Этот эффект лежит в основе зрительной иллюзии, известной как полосы Маха: когда две области, светлая и темная, соприкасаются друг с другом, на краю темной области рядом с границей мы видим узкую сверхтемную полосу, а на краю светлой области – такую же сверхсветлую полосу.
Давайте коротко подведем итог: наша сетчатка покрыта четырьмя основными типами детекторов – с транзиторным on-ответом и off-ответом и с устойчивым on-ответом и off-ответом. Каждый из этих четырех типов подвержен влиянию механизма латерального торможения, который увеличивает силу сигнала у края объекта по сравнению с сигналом в середине однообразного поля. Но, как мы узнаем в главе 4, сетчатка устроена гораздо сложнее – или, как выразились авторы одной научной работы, «оказалась умнее, чем думали ученые»[11]. Выяснить это исследователи смогли не сразу. Но спустя некоторое время технический прогресс позволил заглянуть в мозг и попытаться понять, что мозг делает с информацией, которую получает от сетчатки глаза.
ДЕЛ ЭЙМС: ИЗОЛИРОВАННАЯ СЕТЧАТКА ВИДИТ
Многие Нобелевские премии по биологии были присуждены, по крайней мере частично, за технические инновации. Но на первые полосы попадали далеко не все прорывные технические изобретения и их авторы. Одним из таких новаторов был Дел Эймс, блестящий ученый, щедрой души человек и мой главный учитель.
Эдельберт (Дел) Эймс III родился в Новой Англии в семье, где было много выдающихся личностей, все их заслуги невозможно перечислить. Первый Эдельберт Эймс, дед Дела, был генералом Армии Союза. Сегодня его помнят как просвещенного губернатора штата Миссисипи в период реконструкции Юга после Гражданской войны. Отец Дела, профессор Дартмутского университета, прославился открытием искажающего влияния окружающей среды на восприятие объектов. Возможно, вы слышали о «комнате Эймса» или даже посещали такой аттракцион на ярмарке – это помещение особой конструкции, создающей ложную перспективу, из-за чего внешнему наблюдателю кажется, будто перемещающийся по комнате человек превращается то в карлика, то в гиганта. (Отец Дела также был талантливым скульптором-любителем. Вылепленную им благородную голову индейского вождя – символ банка Shawmut – в те времена можно было увидеть в центре многих городов Новой Англии.)
Дел, высокий худощавый янки, сложением напоминавший Теодора Рузвельта (на внучке которого Дел женился), был заядлым туристом, охотником и рыболовом, смельчаком. Он дожил до 97 лет, но и за год до смерти его часто видели бегущим на лыжах. В студенческие годы Дел входил в лыжную команду Гарвардского университета и затем всю жизнь участвовал в лыжных гонках. В молодости он вместе с университетскими друзьями построил планер. Их безмоторный летательный аппарат разгонялся за счет буксировки автомобилем и после первоначального набора высоты держался в воздухе благодаря восходящим воздушным потокам. Никто из них не умел управлять таким судном, поэтому Дел совершил первый полет и научил остальных. Спустя годы Дел помог своему сыну Дэвиду построить копию того первого планера из легкого деревянного каркаса и перетяжек. Я присутствовал при его запуске. Дэвид бежал по склону холма возле их дома в Конкорде и, когда планер достаточно разогнался, вспрыгнул в него и занял лежачее положение пилота. Машина поднялась на высоту около четырех метров, после чего из-за ошибки пилотирования потеряла скорость – крылья треснули, и планер рухнул на землю.
Вторая мировая война прервала учебу Эймса в Гарвардском университете, где он успел проучиться три года и откуда его в срочном порядке направили в медицинский институт. Впоследствии он даже с некоторой гордостью хвастался тем, что у него нет университетского образования. В Гарварде ему отказались выдать диплом, ограничившись вручением справки о «посещении» лекций. Военные обратили внимание на успехи Дела в изучении медицины, явный научный талант и любовь к холоду и командировали его в Фэрбенкс на Аляске, чтобы заняться исследованиями специфики ведения боевых действий в зимних условиях. Там он пережил один из самых морозных дней за всю историю наблюдений, когда температура воздуха опустилась до –64 ℃. Среди задач, которые военные поставили перед Делом, был поиск наиболее эффективных способов отогревания летчиков и моряков, подвергшихся сильному переохлаждению (как известно, тем же самым занимался печально знаменитый нацистский врач Йозеф Менгеле в концлагерях).
Группа Эймса сделала ряд удивительных открытий. Эксперимент начинался с того, что добровольца погружали в ванну с ледяной водой. Когда температура его тела падала на несколько градусов, испытуемого доставали из воды и согревали. При снижении температуры внутренней части тела до 35 ℃ у человека наступает переохлаждение; понижение еще на несколько градусов ведет к сильной дрожи и болезненному сокращению кровеносных сосудов в руках и ногах, а дальнейшее ее падение грозит смертью. Исследователям нужно было сравнить различные способы согревания жертв гипотермии.
Оказалось, вспоминал Эймс, что некоторые традиционные способы согревания имели серьезные недостатки. Например, если поместить переохладившегося человека в теплую комнату и дать ему стакан бренди, его температура не поднимется, а упадет еще больше. Причина этого парадокса кроется в обычной физике. Даже в очень теплой комнате температура воздуха обычно составляет около 26–27 ℃. При этом температура даже сильно замерзшего человека близка к 35 ℃. Алкоголь расширяет поверхностные кровеносные сосуды, что вызывает приток крови к поверхности кожи, которая соприкасается с более прохладным воздухом (мало кто разогревает комнату до температуры выше 35 ℃). Другими словами, алкоголь стимулирует теплообмен, в результате чего тело замерзшего человека отдает драгоценное тепло окружающей среде, что только усугубляет ситуацию. (Лучший способ – поместить человека под горячий душ.)
После войны Эймс завершил медицинское образование в Колумбийском университете и вернулся в Гарвард, чтобы попробовать себя на исследовательском поприще. Он не сразу занялся нейробиологией, а устроился стажером в лабораторию доктора Бэрда Хейстингса, заведующего кафедрой биологической химии в Гарвардской медицинской школе. Независимый мыслитель, Эймс решил найти способ изолировать нервную ткань для ее изучения. Проще говоря, он хотел вывести мозг за пределы черепной коробки, чтобы его легче было исследовать. Идея была радикальной, и доктор Хейстингс уверенно заявил, что ничего не выйдет, однако Эймс проанализировал метаболизм нейронов и не нашел в нем ничего такого, что помешало бы ему осуществить свою затею.
В те времена в нейробиологии доминировали клинические неврологи, которые были убеждены, что нейроны мозга – чрезвычайно нежны и уязвимы и могут нормально функционировать только в своей привычной среде внутри надежно защищающей их черепной коробки. У медиков были весомые основания так считать. Они знали, что нарушение питания мозга даже на несколько минут ведет к его необратимым повреждениям. При остановке сердца потеря сознания происходит через считаные секунды, и, если не восстановить кровообращение в течение следующих нескольких минут, мозг пациента (и, следовательно, сам пациент) умирает или впадает в вегетативное состояние.
Эймс заинтересовался метаболизмом мозга еще на Аляске. Нейроны действительно требуют очень много энергии – в этом они превосходят практически любую другую ткань организма. Мозг весом всего один – два килограмма потребляет около 20 % общих энергетических ресурсов нашего тела. Вот почему кровоснабжение мозга очень интенсивно; питание его тканей осуществляется через чрезвычайно разветвленную сеть капилляров. Питательные вещества от капилляров к нейронам и продукты клеточной жизнедеятельности в обратном направлении переносятся посредством пассивного транспорта (диффузии). Но диффузия хорошо работает только на очень коротких расстояниях, поэтому мозг опутан частой сетью тончайших капилляров. Студентов-медиков учат тому, что мозговой нейрон может находиться от капилляра на расстоянии не дальше 0,2 мм. Чтобы вам легче было представить: плотность капиллярной сети больше, чем у плетения ткани в обычной простыне.
Эймс задался вопросом, есть ли в нашей центральной нервной системе такое место, где нейроны можно изолировать от окружающих неневральных клеток. В конце концов он нашел такое место – сетчатка. Неспециалисты редко знают о том, что центральная нервная система состоит не только из головного мозга, но и спинного мозга и сетчатки. Эти три структуры имеют сходное эмбриологическое происхождение и образованы из одинаковых видов нейронов и вспомогательных клеток. Все они защищены гематоэнцефалическим барьером – комплексным механизмом, который сохраняет внутри них особую химическую среду, обособленную от остального организма. Большинство нейронов сетчатки и спинного мозга являются полноценными нейронами. Взятые в отдельности нейроны сетчатки (кроме палочек и колбочек) даже для большинства нейробиологов неотличимы от нейронов, находящихся в других частях центральной нервной системы.
Но сетчатка сталкивается с одной проблемой, которой нет у других структур центральной нервной системы: ей нужно воспринимать свет. Если сетчатку пронизать обычной сетью артерий, вен и капилляров, эти сосуды и несомая ими кровь будут преграждать путь свету. В результате мы бы видели мир словно через плотную москитную сетку на окне. Однако эволюция нашла удачный способ решить эту проблему. Сетчатка представляет собой тонкий пласт клеток толщиной обычно не больше 0,3 мм. Благодаря этому бо́льшая его часть находится в пределах досягаемости для диффузии с одной стороны. Сетчатку пронизывает несколько кровеносных сосудов, которые обеспечивают питание самых дальних ее слоев, но основным источником питания служит густая сеть кровеносных сосудов, расположенная снаружи сетчатки в виде сосудистой оболочки.
Эймсу играло на руку и то, что у большинства млекопитающих сетчатка неплотно связана с этим нижележащим слоем и довольно легко от него отделяется. Вот почему отслойка сетчатки – распространенный риск при некоторых заболеваниях или травмах, таких как попадание в глаз хоккейной шайбы или теннисного мяча. Но сама отслаивающаяся сетчатка при этом остается неповрежденной, поэтому, если вовремя вернуть ее на место посредством хирургического вмешательства, пострадавшая сетчатка (и зрение) вскоре восстанавливается.
Эймс разработал формулу искусственного раствора, имитирующего спинномозговую жидкость, которая омывает центральную нервную систему. В ходе своих экспериментов ученый быстро вынимал глаз у находившегося под наркозом животного (его потом усыпляли, пока действие наркоза не прошло), разрезал глаз и осторожно отделял сетчатку, оставляя ее висеть на зрительном нерве. Потом нерв перерезался. Так Эймс получал изолированную сетчатку – тончайшую, почти прозрачную полусферу, бледно-розовую в неподвижном состоянии и становившуюся серебристой при облучении светом. Плавающая в чашке Петри, она была похожа на кусочек намокшей салфетки Kleenex размером с чайную ложку.
Почти по всем параметрам изолированная сетчатка является живой. Она продолжает потреблять кислород и глюкозу, синтезирует новые белки, выделяет продукты обмена веществ. Нейроны сетчатки сохраняют электрическую активность. В течение следующих нескольких лет Эймс и его коллеги доказали, что изолированная сетчатка ведет себя именно так, как предположительно должна вести себя нервная ткань головного мозга. И, что самое главное, она реагирует на свет точно так же, как сетчатка в глазу живого существа.
В последующие годы метод Дела переняли все нейробиологи, и к 1980 г. почти никто не исследовал сетчатку непосредственно на животных. Более того, оказалось, что многие другие образцы нервной ткани тоже могут быть изолированы и существовать вне организма при правильной инкубации. Разработанный Делом специальный инкубационный раствор, известный как «транспортная среда Эймса», теперь продается компанией Sigma-Aldrich, ведущим поставщиком лабораторных химикатов[12]. По моим весьма приблизительным подсчетам, за последние 40 лет они продали около 300 000 л этого раствора, что достаточно для того, чтобы спустить на воду фрегат ВМС США. (Эймс никогда не требовал роялти или каких-либо других выплат за свое изобретение. Впоследствии, когда ему была нужна для экспериментов «транспортная среда Эймса», он за деньги покупал ее у Sigma-Aldrich.)
ЗАВЕСА ПРИОТКРЫВАЕТСЯ
Меня заинтересовала технология Эймса, и по окончании магистратуры я добился, чтобы меня приняли научным сотрудником в его гарвардскую лабораторию. Эксперименты, которые я проводил под руководством Эймса, могли бы служить отличным базовым введением к курсу биологии восприятия. В них не было ничего новомодного, ничего «нобелевского» – просто реальная наука, небольшой, но важный шаг вперед, который открывал путь к новому пониманию.
Мы с Эймсом хотели узнать, как работает нейронная сеть сетчатки – проникнуть внутрь сетчатки и понять, каким образом происходит передача сигналов от внутренних нейронов к ганглионарным клеткам, которые в итоге формируют зрительное сообщение для головного мозга. Чтобы узнать это, мы собирались воздействовать на синапсы нейронов с помощью различных веществ, нацеленных на конкретные типы синапсов. Проще говоря, мы хотели оказать точно выверенное химическое воздействие на систему и посмотреть, как она отреагирует.
Первым шагом на пути к нашей цели было узнать, какие именно нейромедиаторы и каким образом используются в сетчатке. Нейроны сетчатки связаны десятками видов синапсов, и мы хотели детально изучить работу этой системы – простимулировать отдельные синапсы и посмотреть, как меняется выходной сигнал сетчатки. Например, существуют ли конкретные нейромедиаторы, отвечающие за передачу только on-ответов или только off-ответов? Какие нейромедиаторы вовлечены в способность сетчатки воспринимать движущиеся стимулы? Не приведет ли это к открытию механизма, который кажется магическим и посредством которого небольшая группа нейронов в сетчатке определяет направление движения стимула?
Сама схема эксперимента была довольно простой. Глядя через микроскоп, я осторожно опускал микроэлектрод, пока его кончик не касался поверхности сетчатки. Если мне везло, ганглионарная клетка издавала легкий хлопок (мы выявляем активность нейронов, усиливая слабый сигнал, улавливаемый микроэлектродом). Если нет, я осторожно перемещал микроэлектрод влево или вправо, внимательно слушая, когда появится отчетливая последовательность электрических разрядов. Когда клетка была надежно изолирована, я включал примитивный оптический стимулятор с воздушным охлаждением, который излучал на сетчатку точечное пятно света. Во время вспышек света я прислушивался к тому, как реагирует клетка. Изучив параметры ее ответа, я вводил через боковой отвод тестовые реагенты и смотрел, изменилась ли реакция. Все это делалось почти в полной темноте, при тусклом красном освещении подобном свету ночника, чтобы минимизировать непреднамеренное раздражение сетчатки. Звуковым фоном служило шипение воздуха в вентиляции и потрескивание фоновых разрядов – нейронного шума.
И да, забыл сказать, что воздух в комнате был нагрет до температуры тела кролика (37,2 ℃). Когда Дел разрабатывал оригинальный эксперимент, он не знал, какие условия необходимы, чтобы ткань оставалась живой вне организма. Ему показалось логичным предположить, что одно из ключевых требований – держать изолированную сетчатку в таких же температурных условиях, которые существуют в кроличьем глазу. Но сетчатка инкубировалась в проточном растворе, в который вводилась струя кислорода. Так как же гарантировать поддержание стабильной температуры в инкубационной чашке? Всегда дотошный, Дел нашел простое решение: создать среду, в которой всё – сетчатка, растворы, воздух – имело температуру тела кролика, 37,2 ℃. По его заказу в лаборатории построили небольшую «теплую комнату», которая нагревалась внешними обогревателями до любой необходимой температуры. Зимой, когда воздух был сухим, находиться 12 часов подряд в крошечной комнатушке, нагретой до 37,2 ℃, не составляло труда. Куда сложнее нам приходилось летом, когда воздух насыщался влагой. (Когда я создал собственную лабораторию, чуть ли первым делом задался целью найти другой способ контролировать температуру.)
В те времена было известно всего несколько видов нейромедиаторов. Как показал грубый химический анализ, все они присутствовали в сетчатке. Заинтригованные, мы решили использовать эти вещества в качестве нейрональных маркеров, которые помогли бы нам идентифицировать клетки. Исходя из предположения, что разные функциональные типы нейронов должны использовать разные нейромедиаторы, мы решили, что изучение действия этих медиаторов укажет нам на то, какие клетки сетчатки выполняют специализированные функции.
Самым хорошо изученным на тот момент нейромедиатором являлся ацетилхолин. Было установлено, что сетчатка содержит чрезвычайно высокий уровень ацетилхолина: его концентрация здесь выше, чем практически в любой другой структуре нервной системы. Предварительные эксперименты Эймса и Дэниела Поллена показали, что некоторые ганглионарные клетки реагируют на действие ацетилхолина. И поскольку этот медиатор открыли довольно давно, уже было разработано множество препаратов, влияющих на опосредованные ацетилхолином синапсы.
Практически сразу я обнаружил, что многие ганглионарные клетки сетчатки действительно возбуждались под действием ацетилхолина или ацетилхолиноподобных веществ. Они показывали стабильный ответ, возбуждаясь не только ацетилхолином, но и агентами, усиливающими его действие. Другие ганглионарные клетки не реагировали на ацетилхолин. Но выявить в их реакциях / отсутствии реакций какую-либо явную закономерность мне не удалось. Я предположил, что on-клетки должны быть более чувствительны к ацетилхолину, чем off-клетки, но экспериментальные данные не дали этому надежного подтверждения (из-за недостатков, как я теперь понимаю, тогдашней системы классификации ответов).
Тогда я решил зайти с другой стороны и выяснить, какие клетки содержат ацетилхолин. По тем временам это была сложнейшая задача, и, если бы не самоотверженная помощь моего друга Джона Миллса, мастера магической техники вмораживания меченого ацетилхолина, я бы вряд ли сумел это сделать. Наградой за наш кропотливый труд стало единственное открытие, что ацетилхолин содержится в одной небольшой группе амакриновых клеток. (Амакриновые клетки – промежуточные нейроны; модифицирующие возбуждение ганглионарных клеток. Дальше я расскажу о них подробнее). Впоследствии эти клетки получили название «звездчатые», потому что их изящная симметричная форма напомнила Теду Фамильетти, нейроанатому с богатым воображением, фейерверк в виде звезд. Оказалось, что именно эти клетки лежат в основе такой замечательной способности ганглионарных клеток сетчатки, как избирательность в отношении направления.
В те же годы я реализовал еще пару небольших исследовательских проектов, но, чтобы сделать наши главные открытия, нам потребовалось почти семь лет непрерывной работы.
ПУТЬ ВПЕРЕД
Итак, мы установили, что ацетилхолин играет в сетчатке роль нейромедиатора и содержится в небольшой группе амакриновых клеток. Но это был только один нейромедиатор, а нас интересовали и все остальные. Биохимические эксперименты показали, что в сетчатке присутствуют и другие известные кандидаты в нейромедиаторы – например, дофамин, который в головном мозге отвечает за чувство удовлетворения, удовольствия и привязанности. (Нет, это не означает, что сетчатка является частью системы удовольствия – здесь дофамин действует по-другому.) Международная группа ученых во главе с Берндтом Эхингером из Швеции поставила перед собой задачу определить, какие клетки сетчатки содержат другие нейромедиаторы. По мере развития методологий и технологий такие исследования стали намного проще, и я со своей лабораторией присоединился к этим усилиям, хотя и с собственной повесткой.
На мой взгляд, просто составлять список нейромедиаторов сетчатки было скучным делом; гораздо любопытнее было то, что разные нейромедиаторы служили маркерами конкретных типов клеток. В отличие от большинства наших собратьев по цеху, я и горстка других ученых настаивали на том, что нам необходимо знать полные формы различных типов клеток и их реальное количество в сетчатке. Так мы смогли уйти от бессистемного подхода, свойственного классической анатомии. В этом подходе старой школы, который некоторые критики называли коллекционированием бабочек, вы собирали красивые единичные примеры и составляли из них коллекцию – что и становилось вашим исследованием.
Меня же интересовало количество, связи и полные деревья нейронов – разных типов нейронов, которые мы могли идентифицировать благодаря содержанию в них конкретных нейромедиаторов. («Деревом» нейрона (arbor) называется все разветвление его аксонов и дендритов; поскольку эти отростки образуют контакты с другими нейронами, дерево нейрона определяет всю совокупность его возможных связей.) Зная полные структуры и количество разных типов нейронов, мы могли нарисовать нейронную схему сетчатки и таким образом понять, как она функционирует.
Я осознал важность такой методики благодаря одному потрясающему докладу, услышанному мной на конференции по проблемам исследования зрения. Докладчиком был Хайнц Вессле, высокий немец примерно моего возраста, директор Института исследований мозга Макса Планка во Франкфурте. Институты Макса Планка – это научно-исследовательские учреждения, по сути, большие лаборатории, каждую из которых возглавляет один ученый. Они щедро финансируются правительством Германии. Директора Институтов Макса Планка – сливки немецкой науки, а Вессле на тот момент был самым молодым из них.
На лекции, состоявшейся в конференц-отеле рядом с роскошными пляжами западной Флориды, Вессле рассказал о результатах исследования ганглионарных клеток, недавно проведенного им вместе с Брайаном Бойкоттом[13]. Они нашли способ окрашивать два типа ганглионарных клеток сетчатки: крупные ганглионарные клетки, присутствующие в сетчатке в относительно небольшом количестве (они назвали их альфа-клетками), и более мелкие и многочисленные ганглионарные клетки, названные бета-клетками. Затем Вессле вместе с Бойкоттом и его учеником Лео Пайхлем показали, что разная анатомическая форма альфа– и бета-клеток соответствует разной кодировке их визуального ввода. Альфа-клетки были клетками с транзиторными on– и off-ответами; бета-клетки – клетками с устойчивыми on– и off-ответами.
Почему меня так взволновала эта новость? Во-первых, это означало, что уникальная форма клетки свидетельствует о том, что она играет свою конкретную и уникальную роль в функционировании сетчатки. Чем больше мы об этом узнавали, тем больше убеждались, что разные формы всегда означают разные функции в машине сетчатки – подобно разным шестеренкам и колесикам в сложном механизме. Это давало нам возможность двигаться в обратном направлении от формы клеток к нейронным микросхемам и дальше к общей схеме, которая и определяла функцию каждой клетки. Таким образом, изучая форму и количество отдельных деталей, мы могли узнать, как работает этот таинственный механизм – сетчатка глаза, – кодирующий видимую картину мира в набор сигналов для головного мозга.
Второй причиной, почему меня это воодушевило, был уровень определенности, которого добились Вессле и Бойкотт. Проведенное ими анатомическое исследование дало нам не просто красивые бессистемные картинки альфа– и бета-клеток в духе коллекционирования бабочек, но воспроизводимую информацию о популяциях клеток в целом. Это делало открытие стереотипности форм поистине прорывным: аналогично тому, как клены имеют свою типичную форму ветвления, отличающуюся от ветвления дуба, альфа-клетки имели свою специфическую структуру, а бета-клетки – свою. Такое сложно увидеть, когда перед вами всего один экземпляр данного типа клеток – одно кленовое дерево, но когда вы видите всю совокупность клеток данной популяции, их общие черты бросаются в глаза. Немного попрактиковавшись, можно научиться с первого взгляда распознавать альфа– и бета-клетки. И, тогда как Вессле с коллегами открыли всего два типа клеток, было очевидно, что существуют и другие.
Я прилетел на конференцию вместе с двумя гарвардскими друзьями, но, поскольку их интересы лежали в смежных научных областях, они решили пропустить доклад Вессле. Я оставил их в сколоченном из грубых досок баре на набережной под шелестящими кронами пальм, в которых мягко играл легкий бриз из Мексиканского залива. Когда я вернулся, они заканчивали второй кувшин пива. «То, что я только что узнал, навсегда изменит наш подход к изучению нейронных сетей», – сказал я им.
«И что же это такое?» – с нетерпением спросили они.
Я рассказал им об исследовании Вессле и объяснил, что вскоре мы сможем идентифицировать целые популяции клеток и, опираясь на их стереотипные формы, определить их функции. Наконец-то мы сможем прийти к чему-то системному, построенному на надежной количественной и воспроизводимой основе!
Я видел, что они разочарованы. «Анатомия? – думали они. – Ты, наверное, шутишь?!» Но доклад Вессле выкристаллизовал мое мышление: я четко увидел алгоритм, путь вперед, который рано или поздно должен был привести нас к пониманию того, каким образом работает система зрительного восприятия.
Как показало будущее, знание того, как организованы нейроны в сетчатке, – и, в частности, представление об их функциональном разнообразии, – также помогло нам глубже понять другие структуры центральной нервной системы.
4 | Нейроны-призраки
Либо ты, Тиресий, знаешь это наверняка, либо не знаешь вовсе.
ЭЗРА ПАУНД
Тихая революция, произошедшая в нейробиологии в XXI в., была связана с возрождением анатомии. К тому времени некоторые считали анатомию устаревшей наукой, в которой не было места для прорывных открытий. Тем не менее никто не отрицал важности изучения структуры головного мозга. Работа основоположника и покровителя нейробиологии Сантьяго Рамона-и-Кахаля была всецело основана на нейроанатомии. Студенты-медики на протяжении всех последних поколений зубрили названия отделов, ядер и трактов мозга. В широком смысле нейроанатомия, или, как ее теперь иногда называют, структурная нейробиология, говорила нам следующее: мозг – это машина связей и все, что он делает, в конечном итоге сводится к тому, как соединены между собой различные его части.
На рубеже ХХ – XXI вв. ряд технических достижений привел к скачку в понимании анатомии мозга. Первым было значительное улучшение разрешения микроскопов, что было связано с изобретением так называемого конфокального микроскопа (я покажу вам один из них в действии ближе к концу книги). Вторым стало развитие способов визуализации клеточных компонентов. Магические инструменты молекулярной биологии дали нам возможность создавать маркеры даже для самых крошечных частей субклеточного аппарата, а конфокальные микроскопы позволили наблюдать за его работой. Мы получили возможность видеть то, о чем раньше могли лишь мечтать: клетки в движении, плавающие в своей естественной среде; клеточные кластеры, где разные типы клеток светятся в темноте разными цветами, и т. д. Эти достижения позволили нам замахнуться на, как казалось ранее, немыслимое: составить полную опись всех нейронов головного мозга (и сетчатки, в частности), что должно было стать первым шагом на пути к распутыванию его замысловатой системы связей.
ЗАГАДОЧНЫЕ НЕЙРОНЫ
Проведенное Хайнцем Вессле исследование ганглионарных альфа– и бета-клеток, о котором я узнал на конференции во Флориде, показало нейробиологам, что мы можем подойти к изучению сетчатки с другой стороны: сначала составить полный список ее компонентов, а затем попытаться выяснить, какие функции они выполняют. Тем более что к тому времени у нас появились новые замечательные инструменты, которые позволяли это сделать.
Одним из таких инструментов была иммуноцитохимия (ИЦХ). Этот метод, получивший широкое распространение с начала 1990-х гг., позволяет обнаружить присутствие практически любой белковой молекулы внутри клетки или ткани. Если вы когда-нибудь смотрели видео с завораживающими светящимися нейронами, знайте, что их, скорее всего, снимали с использованием иммуноцитохимии. Это довольно простая техника, которая обеспечивает потрясающую визуализацию.
Конечно, не обходится без трудностей и разочарований. Как-то моя лаборатория потратила целый год впустую из-за некачественного коммерческого реактива (в финансовом плане этот неэтичный поставщик обошелся американским налогоплательщикам почти в $300 000). Как бы то ни было, нейробиологи с головой погрузились в ИЦХ-исследования: Харви Картен и Ник Бреча, пионеры этого метода; Джули Санделл сначала в Гарварде, затем в Бостонском университете; Берндт Эхингер в Швеции; Хайнц Вессле и Лео Пайхль в Германии; Дайана Редберн и Стив Мэсси в Техасе и, разумеется, я. Благодаря иммуноцитохимии молодой новозеландский исследователь Дэвид Вэйни нашел свое призвание: он прославился своими потрясающе красивыми снимками, сделанными через микроскоп, так что в конце концов ушел из науки и начал карьеру фотографа.
При наличии подходящих ИЦХ-реагентов этот метод позволял увидеть через флуоресцентный микроскоп все клетки сетчатки, содержавшие конкретную молекулу-мишень. При малом увеличении перед вашим взором представало поле светящихся звезд на темном фоне. При большом увеличении можно было детально рассмотреть форму отдельного нейрона, его тонкие отростки, извивающиеся по сетчатке или ныряющие в глубь нее, его структуру связей с другими клетками. Но как найти вещества-реагенты с избирательным воздействием на конкретные молекулы, которые присутствуют в интересующих нас подтипах нейронов сетчатки? Это делалось (и делается до сих пор) методом научного тыка. Лучшими реагентами были и остаются синаптические нейромедиаторы: дофамин, наш старый знакомый ацетилхолин, серотонин и т. п., каждый из которых присутствует в относительно небольшом наборе нейронов сетчатки. (Разумеется, нейроны содержат намного больше различных молекул, предположительно десятки тысяч. Но большинство из них – особенно те, что отвечают за поддержание клеточной структуры и обеспечение клетки энергией, – присутствуют во многих типах клеток не только в сетчатке, но и в головном мозге и других частях тела. Поэтому для нас такие молекулы бесполезны.)
Итак, опубликовав 20–30 научных работ, наша группа накопила достаточно данных, чтобы составить список из дюжины различных типов клеток сетчатки. Каждый из этих типов клеток окрашивался с высокой степенью надежности, что давало нам возможность четко увидеть всю популяцию клеток этого типа по всей сетчатке отдельно от других нейронов. Мы могли измерить их размер, изучить их форму и структуру связей и сосчитать – что, хотя и звучит банально, лежало в основе настоящей науки, которая уводила нас от коллекционирования бабочек в виде отдельных «типичных» клеток и вела к пониманию общей схемы и, как следствие, того, какую функцию выполняют разные типы клеток в зрительной системе. Например, некоторые типы нейронов были очень малочисленны, но протягивали свои дендриты на большие расстояния по сетчатке. Это говорило о том, что эта популяция не могла быть вовлечена в передачу изображения с высоким разрешением. Низкая плотность клеток означала слишком крупные пиксели: каждая клетка передавала информацию о слишком большой области видимого мира, поэтому изображение, получаемое мозгом, должно было выглядеть состоящим из огромных расплывчатых пятен. И наоборот, некоторые типы крошечных клеток присутствовали в сетчатке в огромных количествах, и им была свойственна высокая плотность. Мы сразу же предположили, что эти клетки образуют канал передачи изображения высокого разрешения от фоторецепторов в мозг, и последующие исследования подтвердили наш вывод.
Таким образом, мы и другие лаборатории увлеченно изучали под микроскопом красивые светящиеся картинки и постепенно начинали понимать, как устроена сетчатка, – пока не столкнулись с проблемой отсутствия реагентов для окраски. Нам удалось найти всего несколько маркерных молекул, способных окрашивать конкретные типы клеток, а все остальное, что мы пробовали, не работало. В комнате остался огромный невидимый слон: бо́льшая часть клеток, которые мы сумели идентифицировать, относилась к редким типам. Поскольку иммуноцитохимический метод позволял выделять сразу целые популяции, мы видели, что большинство этих типов клеток распределено по сетчатке с очень малой плотностью: существовали целые области, где маркерные молекулы не окрашивали ни единой клетки. Если сравнить сетчатку с детской картинкой-раскраской, нам удалось раскрасить всего 20 % ее поверхности, а остальные 80 % оставались белым или, точнее, темным пятном.
Мы были обескуражены. Наше стремление разобраться в устройстве системы зрительной сигнализации ганглионарных клеток, казалось, наткнулось на непреодолимое препятствие: если мы не можем идентифицировать большую часть элементов системы, как мы можем надеяться узнать, каким образом эта система производит свои операции, такие как повышение контрастности, избирательность в отношении направления и т. п.?
Я признаю, что нашим желанием составить полный каталог нейронов сетчатки отчасти двигало простое любопытство. Представьте, что вам подарили старинные часы без инструкции по эксплуатации. Вас заинтересовало их необычное устройство. С функцией маятника все более-менее понятно. Но что делает каждая из этих блестящих латунных шестеренок и прочих деталей? Зачем они нужны? Сама Природа, этот божественный часовщик, дразнила наше любопытство.
Проблема с исследованием сетчатки и остальной части центральной нервной системы была в том, что, будучи окрашены неспецифическими красителями, все нейроны выглядели одинаково. Доступные универсальные красители высвечивали только тела клеток, тогда как именно тонкие нейронные отростки – дендриты, принимающие входные сигналы, и аксоны, посылающие сигналы другим клеткам, – делают каждый тип нейрона особым. Именно по этой причине изучение типов нервных клеток в прошлом страдало от отсутствия системности: нам приходилось работать с отдельными экземплярами, которые удавалось окрасить, и в наших теориях было слишком много места для случайности и догадок.
Мы считали, что в изучении сетчатки мы можем добиться прогресса. В отличие от многих других областей мозга, нам была известна ее функция. У сетчатки есть четко определенное начало и конец; информационные потоки текут через нее в одном направлении; она пространственно компактна – расстояние от фоторецепторов до ганглионарных клеток составляет всего около трети миллиметра. На наш взгляд, было вполне достижимой целью создать карту всех клеток сетчатки. Сегодня такую карту всех нейронов и структуры их связей называют нейромом (neurome) – по аналогии с геномом, совокупностью генов живого организма.
ОХОТА НА ПРИЗРАЧНЫЕ НЕЙРОНЫ
Но как подступиться к этой задаче? Перед нами лежала практически неизведанная территория. Даже об основных классах нейронов сетчатки – фоторецепторах, горизонтальных, биполярных, амакриновых и ганглионарных клетках – на тот момент имелись лишь обрывочные сведения. При использовании обычных красителей эти пять типов клеток выглядели почти одинаково, отличаясь друг от друга немногим больше, чем маленькие овалы на рисунке на следующей странице. Мы знали о существовании этих больших классов клеток и примерно догадывались об их количестве, но как получить более точную информацию обо всех элементах системы? Сетчатка выглядела для нас примерно так, как на этом рисунке: мы могли идентифицировать несколько отдельных клеток (здесь они нарисованы как черные кружки с отростками), а остальные (белые кружки) оставались для нас загадками.

За советом я обратился к Элио Равиоле, старшему сотруднику кафедры нейробиологии Гарвардского университета, магу нейроанатомии. Мой вопрос состоял в следующем: может ли электронная микроскопия (одно из многочисленных искусств, которыми он владел) помочь нам увидеть различия между нейронами? Конечно, ответил он, но это потребует невероятно кропотливого труда: кому-то придется сидеть за специальным резаком (микротомом), делать с образцов сетчатки десятки тысяч ультратонких срезов и затем исследовать их под микроскопом. Поскольку у Элио были дела поважнее, он направил меня к своей итальянской ученице Энрике Стреттои. Энрика оказалась талантливым молодым ученым, абсолютно бескомпромиссным в отношении науки. Вместе с Равиолой они провели потрясающее исследование нейронных связей в сетчатке с использованием электронной микроскопии серийных срезов. Энрика привнесла в нашу команду свои навыки, дисциплину и страсть, а также подсказала ключевую идею, благодаря которой мы сумели достичь нашей цели.
«Нам не нужно гробиться над анализом высокого разрешения, – сказала она. – Зачем возиться с тонкостями строения каждой клетки? Давайте просто идентифицировать клетки по их корневым дефинициям – по путям их отростков к синаптическим слоям сетчатки». В масштабе электронной микроскопии отростки нейронов выглядят огромными. Если на то пошло, заметила Энрика, их можно хорошо разглядеть даже через оптический микроскоп с максимальной разрешающей способностью. В этом случае потребуется гораздо меньше серийных срезов, потому что срезы для оптической микроскопии могут быть в десять раз толще, чем для электронной, и охватывать гораздо более обширные области. Таким образом, тестовые образцы сетчатки, по сути, представляли собой трехмерные сплошные объекты, чью внутреннюю структуру мы собирались изучить на основе двухмерных изображений их срезов (в наши дни цифровой визуализации это не представляло бы большого труда, но в те времена все было намного сложнее). Нашей целью было точно идентифицировать все до единой клетки в тестовых образцах.
Взяв подготовленные нами в Бостоне ткани сетчатки, Энрика вернулась в Пизу и принялась за работу. Она делала бесконечные серии срезов и фотографировала каждый срез под микроскопом. Негативы она отправляла нам в Бостон обычной международной почтой (благословенные технологии цифровой фотографии и электронной почты появились только после окончания этого проекта). Еще одним членом нашей команды была Ребекка Рокхилл, мой лаборант. Ребекка трудилась самоотверженно: когда я попросил ее отпечатать несколько тысяч фотографий, она молча закрылась в темной комнате на пять недель и в конце концов вышла из нее с толстенными пачками глянцевых фотографий размером 21,5 на 22 см, все еще источавших едкий запах фотореактивов.
Сидя за длинным столом, мы перебирали стопки снимков, скрупулезно отслеживая каждую клетку. Процесс происходил так: на фотографии № 1 вы видели множество клеточных тел нейронов – неправильной формы профили, срезанные в разных местах. Вы выбирали любую клетку, после чего брали фотографию № 2 и находили на ней ту же клетку, срезанную немного на другой глубине. Затем вы брали фотографию № 3, снова находили эту клетку и т. д., пока на очередной фотографии не обнаруживали выходящий из тела клетки отросток – аксон или дендрит. Теперь вам нужно было отследить, куда идет этот отросток – вверх к фоторецепторам или вниз к ганглионарным клеткам? Вы находили этот отросток на следующей фотографии и на следующей, прослеживая его траекторию до внутреннего или наружного синаптического слоя. Постепенно отросток становился все тоньше и тоньше и в конце концов исчезал. Разумеется, мы не могли проследить аксоны и дендриты до самых их окончаний, поскольку они становились слишком тонкими, чтобы их можно было зафиксировать на фотографии. Но мы могли проследить их достаточно далеко, чтобы с уверенностью сказать, идут ли они к внутреннему или наружному сетчатому слою.
Определение траекторий всех отростков клетки позволяло нам с высокой степенью надежности идентифицировать ее как биполярную, амакриновую или горизонтальную клетку. Мы опирались на корневые характеристики типов клеток: амакриновая клетка протягивает свои отростки только во внутренний синаптический слой сетчатки; горизонтальная клетка – только в наружный; биполярная клетка – в оба слоя.
Затем мы возвращались к первой фотографии и на теле клетки фломастером писали букву «Б» для биполяров, «А» для амакринов и «Г» для горизонтальных клеток. Если это была первая клетка того или иного типа, мы писали «Б1», «А1» или «Г1», после чего переходили к следующей.
Часть этой колоссальной работы проделал я сам, остальное сделали студенты, проходившие у меня летнюю практику. (Если вы думаете, что я испортил студентам лето и навсегда отбил у них вкус к нейробиологии, то это не так. По меньшей мере двое из них стали ведущими нейробиологами.) Поскольку каждой идентифицированной нами клетке присваивался свой инвентарный номер, который указывался на фотографиях, мы всегда могли вернуться и проверить свои выводы. Таким образом, мы вели предельно строгий учет: идентифицировали каждую клетку в образцах ткани из середины сетчатки и подсчитывали точную долю амакриновых, биполярных и горизонтальных клеток. Мы были абсолютно уверены в точности результатов.
Итак, закончив с этой работой, мы могли задать следующий вопрос: какое количество амакриновых клеток отсутствует в нашем реестре установленных клеточных типов? Мы начали с амакриновых клеток, потому что те были самыми большим классом нейронов внутреннего слоя сетчатки и наименее изученным. Другими словами: каково соотношение всех имеющихся в сетчатке амакриновых клеток и тех типов этих клеток, которые мы уже знаем? Ответ нас шокировал: известные нам типы амакриновых клеток в совокупности составляли всего 24 % от общего числа таких клеток.
Ответ обнадеживал разве что своей четкостью. Как вы помните, целью всего этого начинания, заставлявшей нас считать нейроны до поздней ночи, было понять, как сетчатка обрабатывает информацию: каким образом она формирует сообщения, отправляемые ганглионарными клетками в мозг? Другими словами, как cетчатка запускает первый этап зрительного восприятия? Мы оказались в тупике: 76 % потенциальных входов в ганглионарные клетки оставались для нас невидимыми.
ЭНРИКА СТРЕТТОИ
Энрика Стреттои – директор по исследованиям в Институте нейронаук Национального исследовательского совета Италии. Эта невысокая женщина пленяет своей жизнерадостностью и изысканным стилем. Большую часть времени она ходит на каблуках, и даже если изредка является в лабораторию в джинсах, в ее наряде всегда есть что-то креативное – обычные футболки не для Энрики. В моей памяти навсегда запечатлелась картина: Энрика идет по многолюдной улице в Пизе в разгар лета, на ней элегантный белый льняной пиджак, такая же юбка и жемчужные украшения, ее высокие каблуки легко цокают по неровной брусчатке.
Она родилась, выросла и живет в Пизе – городе, где был основан один из первых университетов средневековой Европы. В детстве она жила на втором этаже над продуктовым магазином, который держала ее мать. Сейчас они с мужем Лукой живут в пригороде в добротном фермерском доме, окруженном прекрасным садом. У них двое дочерей: одна стала врачом, другая учится на врача. В отличие от своей темпераментной матери, ее дочери – спокойные и тихие девушки, по крайней мере когда говорят по-английски.
Энрика – трудоголик. Как настоящая итальянка, по выходным она любит готовить для семьи вкусные блюда, но в рабочие дни приезжает в лабораторию первой и уезжает последней. Она не любит дураков, но вежливо их терпит. График жизни у Энрики такой: на много месяцев уйти с головой в работу, в августе как следует отдохнуть с семьей на море или в итальянских Альпах, а Рождество встретить дома в Пизе. Энрика – набожная католичка. Кроме того, они с Лукой увлекаются пением и поют в местной оперной труппе. Свои электронные письма друзьям она подписывает: «Крепко обнимаю, Энрика».
Один из ее проектов направлен на поиск способов противодействовать развитию слепоты, которая вызывается распространенной группой наследственных заболеваний, в совокупности известных как пигментный ретинит. Причина этих заболеваний в дефектных генах, экспрессируемых в фоторецепторах сетчатки. У человека, унаследовавшего один из таких генов, происходит дегенерация фоторецепторов, и человек теряет зрение – иногда за несколько лет после рождения, иногда за несколько десятилетий.
Энрика задалась вопросом, не может ли сенсорный ввод влиять на течение дегенеративного процесса. Вместе со своими учениками она решила проверить эту гипотезу на мышах с такими же мутациями фоторецепторных генов. Они вырастили одну группу таких мышей в обычных скучных клетках, а экспериментальную группу – в клетках, полных «игрушек» – деревянных блоков, по которым можно было лазить, норок, где можно было прятаться, и колес для бега. К своему удивлению, ученые обнаружили, что сетчатки мышей в такой обогащенной среде деградировали гораздо медленнее. Дальнейшие эксперименты показали, что основной положительный эффект дает беговое колесо – точнее говоря, физическая активность – в сочетании с сенсорной стимуляцией[14].
Как именно это работает, до сих пор неясно, и результаты исследований Энрики не вызвали у научного сообщества большого интереса. В том, что физическая активность полезна, нет ничего нового; все знают, что это – «чудодейственное лекарство», способное замедлить и даже полностью предотвратить развитие практически любых заболеваний от головы до пят. Поскольку исследования Энрики всегда отвечают высочайшим научным стандартам, я убежден, что им можно доверять. И я считаю, что пациенты, страдающие пигментным ретинитом, должны знать, что с помощью физических упражнений они могут замедлить дегенеративный процесс в своей сетчатке. Поверьте, если бы я начал слепнуть от такого заболевания, я бы не ленился дважды в день потеть на беговой дорожке.
ПРОЛИВАЕМ СВЕТ НА АМАКРИНОВЫЕ КЛЕТКИ
Так как же нам установить идентичности этих 76 % таинственных амакриновых клеток? Мы перепробовали весь каталог потенциальных иммунохимических маркеров, но это не дало нам ничего нового. Нам нужно было найти новый подход к изучению нейронов сетчатки, который позволил бы идентифицировать все типы клеток.
Прежде всего мы решили сосредоточиться не на молекулярном окрашивании, а на форме клеток. Причудливые паттерны ветвления нейрональных дендритов и аксонов зачаровывали нейробиологов с момента рождения дисциплины. Некоторое время назад в Массачусетском технологическом институте состоялась художественная выставка, главными экспонатами которой были изысканные зарисовки нейрональных деревьев, сделанные Рамоном-и-Кахалем. Разумеется, некоторые скептики упрямо утверждали, что формы клеток не имеют особого значения, поскольку отражают всего лишь историю развития клеток, а не их функции в зрелом состоянии. Но форма клетки важна по одной неоспоримой причине: у нейрона она отражает его синаптические связи.

На рисунке выше показаны три нейрона сетчатки A, В и С – это вид сбоку, как на поперечном срезе сетчатки. Нейроны A и С – амакриновые клетки, которые протягивают свои отростки только во внутренний слой сетчатки в направлении к ганглионарным клеткам. Обратите внимание, что их дендриты доходят до разных уровней внутреннего синаптического слоя. Это очень важно. Амакриновая клетка С не может образовывать синаптический контакт с ганглионарной клеткой B, потому что их отростки находятся на разных уровнях и не касаются друг друга.
Вторая важная вещь, на которую следовало обратить внимание, – насколько далеко простираются нервные отростки. Амакриновые клетки A и C не могут выполнять одинаковую функцию из-за их разной формы, то есть они должны относиться к разным типам клеток. A – маленькая клетка, C – большая. Как вы помните, горизонтальная протяженность отростков нейрона сетчатки определяет размер его рецептивного поля. Клетки с далеко ветвящимися отростками отвечают за восприятие большой области видимого мира; клетки с короткими отростками – маленькой области. Таким образом, амакриновые клетки А и C выполняют разные зрительные функции, отправляют ганглионарной клетке разные виды сигналов и, следовательно, вносят разный вклад в формирование визуального сообщения, которое ганглионарная клетка передает в мозг.
Вопрос был в том, каким образом в настоящей сетчатке можно увидеть полные формы клеток со всеми их отростками, как это показано на рисунке на предыдущей странице? Тело клетки увидеть довольно легко – это основа клетки, хранилище, где находится ее ДНК и органеллы, отвечающие за производство энергии и поддержание клеточной жизнедеятельности. Но отростки нейрона – аксон и дендриты – почти невидимы: они очень тонкие и тесно переплетены с отростками других нейронов. Даже если бы существовал краситель, способный полностью окрасить дендриты до самых кончиков, идентифицировать все дендриты индивидуального нейрона было бы практически невозможно.
Нам требовалось найти такой метод, который позволил бы нам надежно выделять отдельный нейрон среди всех остальных. Кроме того, метод должен был быть управляемым, чтобы мы могли использовать его для систематической выборки популяций амакриновых клеток. Техника, на которой мы в итоге остановились, называлась фотозаполнением (photofilling). Сначала сетчатка погружалась в раствор со светочувствительными молекулами, которые посредством диффузии проникали во все нейроны. Затем мы фокусировали крошечное – площадью меньше нейрона – пятно света на случайно выбранной амакриновой клетке. В ответ на мощный световой стимул в клетке запускалась цепь реакций, в результате которой флуоресцентные молекулы распространялись по всему внутреннему пространству целевого нейрона – и высвечивали его на фоне миллионов его нефлуоресцирующих собратьев.
Трудностей, разумеется, хватало. Например, флуоресцентную клетку нельзя было фотографировать обычным способом, потому что используемый в этом случае свет вызывал реакцию флуоресценции во всех окружающих клетках. Мы решили эту проблему, купив самый высокочувствительный (и астрономически дорогой) цифровой фотоаппарат, позволяющий делать снимки менее чем за одну десятую долю секунды – прежде чем флуоресцентная реакция успевала распространиться вокруг. Также выяснилось, что этот метод работал лучше с маленькими клетками, чем с большими. Как бы то ни было, по мере практики наш оператор Маргарет Макнил, опытный постдок, овладела этой техникой почти в совершенстве. Когда она нацеливала луч света на случайно выбранную клетку, ей удавалось зафиксировать изображение ее дендритного дерева в 94 % случаев. Несколько сотен таких фотографий обеспечили нам довольно-таки репрезентативную выборку всей популяции амакриновых клеток.
Помните вопрос, с которого мы начали это исследование? Если 24 % всех амакриновых клеток, которые мы идентифицировали, относятся к специфическим редким типам таких клеток, то каковы тогда обычные амакриновые клетки? К нашему большому удивлению, ответ заключался в том, что обычных амакриновых клеток не существует.
Что это значит? Мы ожидали обнаружить среди амакриновых клеток несколько основных крупных групп, дополненных малочисленными группами специализированных клеток. Но вместо этого оказалось, что амакриновые клетки довольно равномерно распределены среди разнообразного набора клеточных типов. А это предполагало, что все они играют одинаково важную роль в обработке визуальной информации. Наш вывод, который мы (с небольшими трудностями) опубликовали в ведущем научном журнале, заключался в том, что в сетчатке существует 29 различных типов амакриновых клеток, каждый из которых выполняет свою специфическую функцию по обработке визуального изображения.
Почему это короткое заключение заслуживает особого внимания? Оказалось, в нем крылся важный ключ к разгадке того, как работает сетчатка. Зачем сетчатке нужны целых 29 типов амакриновых клеток? Ответ напрашивался сам собой: в сетчатке происходит гораздо больше обработки информации, чем считалось раньше. Амакриновые клетки генерируют основной выход для ганглионарных клеток, которые являются последним звеном цепи перед отправкой зрительных сообщений в головной мозг. Если амакриновые клетки так разнообразны, значит, сообщения должны быть такими же разнообразными. Это был важный шаг вперед к пониманию того, как работает зрительное восприятие.
НЕЙРОНЫ-ПРИЗРАКИ (ПРОДОЛЖЕНИЕ): БИПОЛЯРНЫЕ КЛЕТКИ
Пока мы занимались амакриновыми клетками, наши коллеги изучали другие компоненты микросхемы сетчатки. Главным пробелом в нашей базе знаний были биполярные клетки. Как вы помните, биполярная клетка получает синаптический вход от нескольких фоторецепторных клеток и передает выходной сигнал через амакриновую клетку в ганглионарную. Биполяры являются важнейшим элементом сетчатки. Если удалить из сетчатки все амакриновые клетки, после некоторой перестройки она все равно сможет функционировать благодаря ганглионарным клеткам с транзиторным и устойчивым ответами, хотя изображение будет не таким контрастным и пропадет избирательность в отношении направления. Другими словами, человек без амакриновых клеток все равно будет видеть, но его зрение окажется расплывчатым и замедленным. Если убрать биполярные клетки, сетчатка сможет сообщать мозгу только о наступлении дня и ночи благодаря примитивной функции, выполняемой подгруппой органически светочувствительных ганглионарных клеток.

Все вы слышали о научных прорывах, когда новое открытие или идея внезапно, подобно молнии среди ясного неба, меняли целые области науки. Но гораздо чаще наука движется вперед путем постепенного приращения: по мере накопления данных возможность превращается в вероятность, которая в конце концов трансформируется в факт. Так было и в случае с нашим пониманием биполярных клеток.
Первые системные исследования с регистрацией электрической активности биполярных клеток, проведенные Акимити Канеко, Фрэнком Верблином и Джоном Даулингом, показали существование четырех типов клеток: с транзиторными on– и off-ответами и устойчивыми on– и off-ответами. Казалось логичным предположить, что эти четыре физических типа биполярных клеток соответствуют четырем аналогичным типам ганглионарных клеток.
Но, как и в случае амакриновых клеток, имелись весомые основания подозревать, что это далеко не все разновидности биполярных клеток, которые существуют в сетчатке. В середине 1990-х гг. исследованием биполярных клеток занимались четыре – пять лабораторий, и по их оценкам количество типов таких клеток варьировалось от четырех до девяти. Мы с моей лабораторией присоединились к этой работе довольно поздно с тайной надеждой на то, что накопленная другими база знаний позволит нам понять организацию биполярных клеток. Но, как оказалось, эта база знаний включала в себя слишком много несистематических данных в духе коллекционирования бабочек: небольшая выборка клеток здесь, небольшая выборка там. Поэтому наш набор вопросов был совершенно иным. Во-первых, существуют ли типы биполярных клеток, которые не удалось обнаружить с помощью ранее использованных методов окрашивания? Во-вторых, существует ли среди биполярных клеток разделение на основные, доминирующие группы и малочисленные группы вспомогательных клеток? Или же все типы биполярных клеток более-менее равны?
Чтобы ответить на вопрос, мы объединились с Элио Равиолой. Некоторое время назад он провел потрясающую серию экспериментов с окрашиванием клеток сетчатки, но эти снимки пылились в нижнем ящике его рабочего стола[15]. Один из его студентов провел предварительный анализ, но Элио решил не продолжать работу, потому что, будучи перфекционистом, остро осознавал несовершенство оборудования. В частности, он считал, что ему удалось окрасить далеко не все биполярные клетки.
У нашей лаборатории имелось два важных преимущества. Во-первых, метод фотозаполнения, который позволял получить надежную и всеобъемлющую выборку биполярных клеток. Во-вторых, у нас была Маргарет Макнил, которая к тому времени стала мастером трехмерной визуализации нейронов. Сделанные ею снимки были нашей тайной гордостью. Мы, нейроанатомы, любим красивые фотографии нейронов – нам кажется, что в них кроется что-то мистическое, открывающаяся нашему взору частица Истины.
Наконец, у нас имелся еще один ценнейший набор данных для идентификации клеток: снимки биполярных клеток, в которые были микроинъектированы маркерные молекулы после того, как были изучены их электрические ответы. Эта работа была проделана нашим другом Рэем Дашё из Алабамского университета. Знание того, как клетки реагируют на свет, было важным дополнением, поскольку реакции клеток оказались такими же разнообразными и характерными, как и их формы. Каждый из трех методов – окрашивание, фотозаполнение и микроинъекции – имел свою специфику, и мы надеялись, что ни одному типу клеток не удастся ускользнуть от всех трех детекторов. Итак, объединив все три вида данных, мы с уверенностью пришли к выводу, что существует целых 13 типов биполярных клеток. Вот они, нарисованные рукой Элио:

Этот рисунок подчеркивает определяющую особенность форм биполярных клеток – глубину их аксонных деревьев. Как и другие лаборатории, мы обнаружили, что главная отличительная черта биполярных клеток – конкретный уровень в синаптическом слое сетчатки, на котором разветвляются их аксоны. Как вы видели на рисунке амакриновых клеток, глубина погружения во внутренний синаптический слой сетчатки влияет на то, с какими типами других игроков – амакриновых и ганглионарных клеток – контактирует эта конкретная биполярная клетка.
Мы также обнаружили, что не существует какого-либо основного, преобладающего типа биполярных клеток. Как и амакриновые клетки, биполяры оказались более-менее равномерно разделены на несколько разных групп. Это означало, что сетчатка имеет примерно 13 параллельных путей от фоторецепторных клеток к внутреннему слою, где они подходят к примерно 29 типам амакриновых клеток и далее идут к, предположительно, еще большему числу типов ганглионарных клеток, которые окончательно кодируют визуальное сообщение и передают его в головной мозг.
Некоторое время спустя, когда были изобретены более совершенные красители, группа Хайнца Вессле провела исследование по идентификации биполярных клеток. Продемонстрировав высший пилотаж в технике иммуноокрашивания, они получили настолько однозначные и детальные данные, что суммирование количества отдельных типов клеток в точности дало общее число биполярных клеток сетчатки (установленное нами с Энрикой). Вессле и его коллеги пришли к выводу, что «все основные типы биполярных клеток сетчатки мыши идентифицированы и каталог из 11 типов колбочковых биполярных клеток и одного типа палочковых биполярных клеток можно считать полным»[16]. Новые исследования, проведенные с использованием самых современных техник электронной микроскопии и мощных молекулярно-генетических маркеров, внесли лишь незначительные изменения и дополнения в каталоги биполярных клеток, составленные Макнил и лабораторией Вессле. Общее количество идентифицированных типов варьируется от 12 до 15 в зависимости от используемых критериев.
Биполярные клетки составляют основу сетчатки. Дюжина типов биполяров представляют собой «примитивы» зрения. На более поздних этапах эволюции сетчатки и мозга сигналы, генерируемые биполярными клетками, стали собираться в различные комбинации, модифицироваться, усиливаться или игнорироваться. Но ни сетчатка, ни мозг не могут выйти за пределы ограничений, установленных биполярными клетками как базовыми элементами зрительного восприятия.
БРАЙАН БОЙКОТТ
Самым влиятельным исследователем сетчатки в конце XX в. был Брайан Бойкотт, член Королевского общества без ученой степени.
Впервые я встретился с ним в его лаборатории в Королевском колледже на Друри-лейн в Лондоне[17]. Это было пыльное, необжитое на вид помещение с деревянными шкафами, забитыми старыми лабораторными журналами и оборудованием. Бойкотт тогда уже был знаменитым ученым. Он возглавлял отделение биофизики в Королевском колледже Лондона, где в свое время никому не известная Розалинда Франклин получила необычайно четкие дифракционные рентгенограммы ДНК, благодаря которым будущие нобелевские лауреаты Уотсон и Крик смогли сделать важные открытия, касающиеся структуры ДНК. Бойкотт носил простую рубашку без галстука и старомодные брюки, над которыми нависало брюшко. Он курил сигареты без фильтра. Я был нервным, амбициозным молодым ученым, только начинавшим приобретать известность за пределами своего университета. Мы сидели лицом к лицу на лабораторных табуретах. Брайан без тени снисходительности или скептицизма расспрашивал меня о моих экспериментах, новости о которых доходили до него по сарафанному радио. Мы проговорили до конца рабочего дня. Это была наша первая из длинной череды столь же затяжных, сопровождавшихся бесконечным курением бесед.
Брайан Бойкотт родился в городке Кройдон, Англия, зимой 1924 г. Когда ему исполнилось семь лет, мать вместе с ним сбежала от мужа-алкоголика. Так в разгар Великой депрессии они остались без финансовой поддержки. Несколько месяцев они жили у друзей, пока матери в конце концов не удалось найти низкооплачиваемую работу и снять комнату.
К счастью, непутевый отец Брайана некогда был франкмасоном, что дало мальчику право на поступление в одну из масонских школ. Это была школа-интернат классического британского образца, где учеников обеспечивали жильем, питанием и даже одеждой. Начиная с восьми лет все детство и юность с перерывом на короткие каникулы Брайан провел в масонской школе.
Хотя трудности Брайана далеко не исчерпывались стандартным набором детских болезней, он не чувствовал себя несчастным в школе-интернате. Его успехи в учебе были невелики. Выпускные экзамены по французскому и математике Брайан провалил, по химии и физике едва наскреб на «удовлетворительно». Отвергнутый Кембриджским университетом, он поступил в Бербек-колледж, филиал Лондонского университета с вечерним обучением, где «народ из рабочего класса по вечерам получал высшее образование». Но вскоре разразилась Вторая мировая война; во время «Лондонского блица» в здание Бербек-колледжа попала зажигательная бомба, поэтому занятия проводились в помещениях уцелевшего первого этажа, которые сверху покрыли обычным гофрированным железом. Студентов дневного отделения Лондонского университета эвакуировали в Северный Уэльс, где они продолжили учебу в комфорте и безопасности, тогда как Бойкотту и его сокурсникам пришлось получать знания под жестяной крышей, которая летом раскалялась под солнцем и оглушительно грохотала под дождем.
Чтобы зарабатывать себе на жизнь, Бойкотт устроился техником в виварий Национального института медицинских исследований, где в его обязанности наряду с прочим входила чистка клеток. Вероятно, он хорошо зарекомендовал себя на этой грязной работе, потому что вскоре ему предложили более интересную должность лаборанта в физиологической лаборатории.
До предыдущего года эта лаборатория была вотчиной сэра Генри Дейла, пионера синаптической биологии, и сохранила заведенные им порядки и дисциплину. Бойкотт проработал там четыре года, по вечерам изучая биологию в колледже. Для начинающего биолога это был фантастический опыт. В виварии он работал плечом к плечу с людьми из рабочего класса, которых уважал и любил. В лаборатории Дейла – с представителями научной элиты. В те времена исследовательские группы были небольшими, и даже у знаменитого на весь мир Дейла группа насчитывала не больше 15 исследователей и лаборантов. По-видимому, даровитый молодой лаборант стал всеобщим любимцем, потому что вскоре ему разрешили самостоятельно проводить эксперименты. Один из таких экспериментов состоял в том, что собак ставили на качающуюся платформу и держали их там, пока у них не начиналась рвота. Это исследование было заказано Королевскими военно-воздушными силами, которые хотели узнать биологическую подоплеку морской болезни. Брайан рассказал мне историю об одной умной собаке, которую начинало тошнить при одном виде платформы, из-за чего в тот день она становилась непригодной для эксперимента. Возможно, именно поэтому нейробиологические основы павловского условного рефлекса всегда вызывали у Бойкотта живой интерес.
Позже он писал о большом значении этого этапа его жизни, когда он сформировал свои исследовательские интересы, сдружился с широким кругом ученых и даже написал первую научную статью. В ней был представлен новый метод измерения количества углекислого газа, накапливаемого в ребризере – подводном дыхательном аппарате с повторным использованием выдыхаемого воздуха. Хотя война к тому времени подошла к концу, к сожалению, военные поставили на статье гриф «секретно» и запретили ее публиковать.
Как бы то ни было, Бойкотт приобрел некоторую известность в научных кругах, и после окончания Бербек-колледжа ему предложили место младшего преподавателя на факультете зоологии в Университетском колледже. Он преподавал студентам лабораторный раздел вводного курса (сейчас эту черную работу обычно возлагают на ассистентов кафедры или аспирантов). Он также записался в аспирантуру, но отложил работу над диссертацией, когда получил предложение войти в исследовательскую группу зоолога с мировым именем Дж. З. Янга, работавшего в лаборатории морской биологии в итальянском Неаполе. Море было щедрым источником животных, на которых зоологи могли изучать общие принципы, а Бойкотта, в частности, интересовало, можно ли узнать что-то новое о механизме обучения, используя мозг осьминога.
Проведенное группой Янга исследование нейронных основ обучения вызвало своего рода ажиотаж. О нем много писали не только в различных научных журналах, но и в популярной прессе (еще бы, осьминоги способны учиться!). К сожалению, звездного босса Брайана больше заботила публикация очередной статьи, чем диссертация его сотрудника. В результате Бойкотт так и не написал диссертацию, хотя к тому времени уже заработал себе солидную научную репутацию. Британское научное ведомство решило, что докторская степень не так уж важна для настоящего ученого, и повысило Бойкотта до профессорского звания. Впоследствии он с гордостью поправлял каждого, кто называл его доктором Бойкоттом, – мол, он «просто профессор Бойкотт».
По возвращению из Неаполя в Великобританию ученый заинтересовался другими темами, связанными с обучением, в частности изменениями, которые происходят в мозге у представителей семейства беличьих, когда те впадают в спячку. Отчасти по этой причине он на один семестр отправился преподавать в Гарвард, где найти этих пушистых грызунов для опытов было куда легче, чем в Лондоне.
В Гарварде он познакомился с Джоном Даулингом, и партнерство между ними привело к важному достижению. На тот момент Даулинг как раз занялся изучением сетчатки млекопитающих с помощью электронной микроскопии. Но электронная микроскопия дает слишком большое увеличение. В результате Даулинг обнаружил в сетчатке любопытную организацию синаптических связей, но из-за чрезмерно увеличенного изображения никак не мог понять, каким клеткам они принадлежат. Бойкотт был мастером окрашивания целых клеток и, изучая осьминогов, научился искусно распутывать нейронные сети. Бойкотт и Даулинг быстро осознали, что прекрасно дополняют друг друга. Вместе они провели фундаментальное исследование нейронных связей в сетчатке, которое заложило основу для ее понимания.
Бойкотт и сам обладал уникальным видением: он мог увидеть целиком всю картину и с точностью лазера определить набор ключевых вопросов для последующего движения вперед. Его собственная исследовательская работа, которой он занимался с помощью всего одного лаборанта, касалась микроструктуры сетчатки. Но, пожалуй, его главным вкладом было умение донести свою широту мышления до нас, молодых ученых. Несколько месяцев в году он проводил в лаборатории Хайнца Вессле во Франкфурте в качестве консультанта, критика, серого кардинала и источника кипучей энергии. Вессле, выдающийся ученый и грамотный администратор, предоставлял в его распоряжение все ресурсы Института Макса Планка. Брайан генерировал общие идеи, сотрудники Вессле корпели над частностями. Несмотря на разные поколения, Бойкотт и Вессле были людьми одного типа: великодушными, высоконравственными, преданными своему делу.
Предпочитая вести простой образ жизни, Бойкотт тем не менее не был пуританином. Он интересовался мировыми событиями, политикой, любил поесть и выпить. В любой компании он первым предлагал отправиться в паб. Придерживаясь высочайших научных стандартов и обладая очень критическим мышлением, он на равных заводил разговор с кем угодно – от дворника и постдока до именитого профессора – о чем угодно, будь то случайная тема, политика, мудреная научная теория и т. д. Если вам было что сказать, он уважительно вас выслушивал. Если же ваши стандарты, по его мнению, не соответствовали его высокой планке, он оставался дружелюбным, но не придавал вашему мнению большого значения. Крупных научных сборищ он избегал, насмешливо называя их собранием овец. К публичным выступлениям почти не готовился, поэтому его речи нередко напоминали сбивчивые размышления вслух, будучи полной противоположностью современным презентациям в стиле TED, тщательно подготовленным и отлично проиллюстрированным.
Брайан любил общаться с молодыми учеными, чьи исследования вызывали у него интерес. Во время поездок в США он часто приезжал в мой маленький домик под Бостоном. Мы допоздна засиживались на заднем крыльце, обсуждая популяции клеток сетчатки, сплетничая о друзьях и потягивая бурбон. После смерти Брайана ученые, занимающиеся исследованиями сетчатки, учредили в его память награду за научные достижения – ее вручают раз в два года, когда все сообщество собирается в горах Вермонта на научное биеннале. Стать лауреатом премии Бойкотта – большая честь, поскольку Брайана все уважали и любили. Победитель получает сертификат и бутылку односолодового виски.
5 | Что глаз сообщает мозгу?
Водной реализации кодировщик, использующий первый набор выученных примитивов визуального знания, исключает визуальную информацию из изображения до сжатия. После распаковки декодировщик, использующий автономно выученный набор примитивов визуального знания, восстанавливает исключенную визуальную информацию и синтезирует изображение.
ИЗ ЗАЯВКИ НА ПОЛУЧЕНИЕ ПАТЕНТА США НА АЛГОРИТМ ВИЗУАЛЬНОГО РАСПОЗНАВАНИЯ
Невозможно познать всю реальность, но вполне возможно понять ее.
БРАЙАН ТОМПСОН. ДЗЕН-МЫШЛЕНИЕ
Теперь, когда мы познакомились с основными участниками работы сетчатки, мы можем перейти к следующему этапу – к обретению понимания того, как они передают зрительную информацию остальной нервной системе.
Вы помните главный принцип: разрешающая способность зрения определяется плотностью мозаики ганглионарных клеток на сетчатке – аналогично тому, как плотность пикселей на экране определяет его разрешение. Проще говоря, чем плотнее расположены ганглионарные клетки, тем острее зрение.
Вы также знаете некоторые базовые принципы того, как сетчатка воспринимает зрительные раздражители: одни ганглионарные клетки реагируют преимущественно на действие светового стимула, другие на его ослабление, при этом некоторые из них генерируют короткий транзиторный ответ, другие – устойчивый ответ на протяжении всего времени действия стимула.
Разумеется, этим знания современной науки о ганглионарных клетках не исчерпываются. Недавние исследования показали, что у большинства млекопитающих насчитывается более 30 различных типов ганглионарных клеток, каждый из которых отвечает за восприятие определенных аспектов зрительного раздражителя. Далее я расскажу вам о некоторых из этих типов. Но детали не так важны, как понимание главного, а именно того, что ганглионарные клетки формируют окончательные сообщения, которые сетчатка посылает в головной мозг. Это единственные сообщения, которые мозг получает от глаза, поэтому они полностью определяют то, что наш мозг знает о видимом мире и каким он его «видит».
«УМНЫЕ» ГАНГЛИОНАРНЫЕ КЛЕТКИ
Помимо уже известных нам четырех типов ганглионарных клеток сетчатки (с транзиторными или устойчивыми on– или off-ответами) существуют и другие. Среди «умных» типов ганглионарных клеток лучше всего изучены клетки с избирательной чувствительностью к направлению, которые реагируют на движение стимула в одном направлении и не реагируют на движение того же стимула в противоположном направлении. Другими словами, такая клетка реагирует на направление движения независимо от конкретного визуального объекта. Ей неважно, что именно движется через ее рецептивное поле – светлая часть объекта или темная (что с точки зрения физики является совершенно другим стимулом), точечное световое пятно или крупный объект, – если этот раздражитель движется, скажем, слева направо, клетка возбуждается. На рисунке ниже пунктирной линией обозначено рецептивное поле клетки; маленьким кружком внутри него – зрительный стимул. Клетка реагирует на движение стимула, чей размер меньше ее рецептивного поля, независимо от того, в какой части поля происходит это движение. В основе этой способности лежит изобретательный нейронный механизм. Я не буду здесь вдаваться в детали, но, когда наши немецкие коллеги в 2015 г. разрешили эту загадку, мы все праздновали победу.

Нам известно, что этот тип клеток, в частности, помогает контролировать положение глаз во время движения. Подумайте, что происходит, когда вы смотрите в окно быстро движущегося поезда или автомобиля. Видимая картина стремительно уносится назад, и, если бы мы были способны удерживать глаза в неподвижном состоянии, мы бы видели размытое изображение. На самом деле мы не можем сознательно помешать глазам следить за движущимся объектом: они перемещаются назад вслед за ним, затем перепрыгивают вперед, навстречу ему. Если вы сомневаетесь в этом, попросите кого-нибудь последить за вашими глазами, когда вы смотрите в боковое окно мчащегося автомобиля.
Важную роль в этом рефлексе играют ганглионарные клетки сетчатки, избирательные в отношении направления. Когда изображение объекта перемещается по сетчатке, эти чувствительные к направлению нейроны возбуждаются и сообщают мозгу, что изображение движется и в каком именно направлении. Эта информация передается в соответствующий отдел мозга, который посылает ответные команды глазным мышцам, приказывая им двигать глазами определенным образом, чтобы стабилизировать изображение на сетчатке.
Этот рефлекс важен не только при езде в поезде или автомобиле. С аналогичной проблемой мы сталкивается и при ходьбе, причем характер движения здесь намного сложнее: когда мы идем, то, по сути, перепрыгиваем из точки в точку. Нашим глазам приходится приспосабливаться к этой своеобразной траектории, и они делают это благодаря избирательным к направлению ганглионарным клеткам, которые помогают компенсировать эти сложные движения и стабилизировать воспринимаемое изображение во время ходьбы. Попросите кого-нибудь подвигать страницу с крупным шрифтом из стороны в сторону перед вашими глазами в то время, как вы стараетесь неподвижно смотреть прямо перед собой, – вот так выглядел бы мир без механизмов стабилизации изображения.
Еще один тип «умных» ганглионарных клеток сетчатки – детекторы локальных контуров (local edge detector). Эти клетки реагируют на очень медленное движение крошечного пятна в пределах своего рецептивного поля. Крупные раздражители их не возбуждают – на самом деле их не привлекает практически ничего из того, что можно увидеть на уровне земли. Уильям Левик, открывший этот тип клеток в сетчатке кроликов, предположил, что это может быть эволюционной адаптацией зрения животного, которое является объектом охоты. Например, именно такой крошечный движущийся стимул генерирует на сетчатке ястреб, медленно кружащий высоко в небе. Этот тип ганглионарных клеток был обнаружен в большом количестве у наземных грызунов, в том числе у мышей, которым также необходимо следить за небом, опасаясь угрозы нападения.
Впервые такие нейроны с избирательным восприятием были найдены в сетчатке лягушек. Тогда исследователи – вполне логично – сочли их детектором насекомых. Но на лягушек также охотятся ястребы. Так какова же на самом деле роль этих клеток – обнаруживать хищников или мелкую добычу – насекомых? Ответ таков, что пока у нас нет окончательного ответа. Чтобы точно определить предназначение этих и других типов клеток, нам прежде всего необходимо понять процесс зрительного восприятия в целом – как мозг вычисляет и формирует картину видимого мира на основе всех поступающих в него входных данных. Пока же нам придется довольствоваться антропоцентричными предположениями о поведении этих клеток, которые, впрочем, напоминают нам об одной важной вещи: эволюция создала эти причудливые нейроны с одной главной целью – помочь данному живому виду выжить в его конкретном видимом мире.
Последний пример – тип клеток со скучным названием «подавляемые контрастом клетки». Вся их реакция заключается в том, что, когда в их рецептивное поле попадает край объекта, они замолкают. Только подумайте: край – с его выраженным перепадом освещенности – не вызывает у них никакой реакции! Причем эти клетки не просто замолкают, но сохраняют молчание, пока край находится в их поле зрения. Их отличительной особенностью, также описанной Левиком, является высокий уровень спонтанной активности в отсутствие стимуляции. В результате, когда клетки вдруг замолкают, этот сигнал привлекает внимание экспериментатора – и предположительно мозга.
Я упомянул о подавляемых контрастом клетках, потому что их полезность для животных пока совершенно непонятна. И это не единственная загадка. Как я уже говорил, есть множество других типов клеток, о роли которых в зрительном восприятии мы пока можем только гадать. Мы знаем, что эти типы клеток существуют: они имеют характерную форму, равномерно покрывают сетчатку и кодируются разными наборами генов. Но у них нет такой простой и очевидной функции, как, например, функция определения движения. Так что в разнообразной мозаике ганглионарных клеток сетчатки в настоящее время остается еще много белых пятен[18].
КАК МОЗАИКА ГАНГЛИОНАРНЫХ КЛЕТОК ВИДИТ МИР
До сих пор мы говорили в основном об отдельных типах ганглионарных клеток, каждый из которых посылает в мозг свой зрительный сигнал. Как известно, плотность ганглионарных клеток определяет остроту зрения. В человеческой сетчатке насчитывается около миллиона ганглионарных клеток. Как все это разнообразие типов клеток соединяется в единую машину, чтобы обеспечивать эффективное восприятие внешнего мира?
Давайте начнем с простого: с одного типа ганглионарных клеток, который сообщает мозгу об одном важном аспекте видимого мира. С одной стороны, эти клетки должны располагаться по всей сетчатке, чтобы избежать пробелов в восприятии данного конкретного аспекта. С другой стороны, избыток нейронного оборудования – это плохо. Поэтому сетчатка стремится использовать для выполнения каждой задачи минимально необходимое количество ганглионарных клеток каждого типа, и они покрывают поверхность сетчатки так, как показано на рисунке ниже.
Теперь давайте представим сетчатку, содержащую не один, а три типа ганглионарных клеток. На рисунке ниже каждый тип обозначен своим цветом.

Предположим, что один из этих типов – стандартные on-клетки; второй – клетки, избирательные в отношении направления; третий – клетки, подавляемые контрастом. На рисунке выше показано, что поверхность сетчатки «вымощена» всеми тремя типами клеток, но ни один не покрывает всю поверхность полностью. Два из трех типов клеток отсутствуют на каждом отдельно взятом участке. Если бы наша сетчатка действительно была организована таким образом, мы бы страдали пробелами в зрительном восприятии – точнее говоря, пробелами в восприятии конкретного видимого свойства, за которое отвечает отсутствующий тип клеток. Например, если бы на этом участке отсутствовали клетки, избирательные в отношении направления, мы бы не могли видеть движущиеся стимулы. (Есть несчастные люди, которые рождаются без этого типа нейронов, из-за чего их глаза постоянно совершают быстрые осцилляторные движения из стороны в сторону.)
В действительности каждый из трех типов ганглионарных клеток покрывает сетчатку независимо от других типов, как показано на рисунке ниже. Мозаика каждого типа клеток накладывается на все остальные. Таким образом, если ткнуть иголкой в сетчатку в любой точке, то в нашем примере вы попадете в рецептивные поля всех трех видов ганглионарных клеток.

Видимый мир проецируется как изображение на сетчатку глаза, и это приводит нас к важному факту о механизме зрительного восприятия: о каждой точке видимого мира мозг получает информацию примерно от 30 различных анализаторов (ганглионарных клеток), каждый из которых сообщает об определенном свойстве этого мира в данной точке.
На рисунке на следующей странице перечислены некоторые из параметров конкретной точки изображения, о которых сетчатка сообщает головному мозгу (точка расположена на плече баскетболиста и обозначена черным кружком). Итак, разные типы нейронов сетчатки посылают в мозг разные сигналы. Например, один набор нейронов сигнализирует мозгу о том, что данная часть видимого объекта движется вправо, влево, вверх или вниз. Другой набор нейронов сообщает о ее цветовом спектре. Детекторы локальных краев, наоборот, посылают слабый сигнал, так как в этой части изображения нет границ и локальных контуров и они видят более-менее однородное поле. Наконец, вопросительными знаками обозначены сигналы остальных типов ганглионарных клеток, о существовании которых мы знаем благодаря их специфической анатомии и экспрессии генов, но нам ничего не известно о том, о каких именно проявлениях видимого мира они сообщают мозгу.
Другой способ проиллюстрировать ту же идею – вообразить, каким бы мы видели мир, если бы в нашей сетчатке отсутствовали те или иные типы ганглионарных клеток. На рисунке на следующей странице представлены два таких изображения (сделанных с помощью Adobe Photoshop): без чувствительных к краям ганглионарных клеток видимый нами мир был бы таким же расплывчатым и нечетким, как изображение президента Линкольна слева; и наоборот, если бы видимая нами картина формировалась только чувствительными к краям нейронами, она была бы слишком резкой, лишенной оттенков, как изображение в центре. Совместная работа этого и других типов клеток дает нам привычное изображение с выраженными контурами и нюансами, как портрет Линкольна справа.

Тридцать разных параметров в каждой точке – это огромное количество информации, которая, безусловно, очень полезна, – но как мозгу переработать весь этот объем? Как все эти разрозненные сигналы, на которые скрупулезно раскладывается исходное изображение, собираются в единую картину видимого мира? То, что субъективно кажется нам целостным изображением, в действительности представляет собой совокупность множества различных репрезентаций. Как эти отдельные изображения вновь объединяются в единое целое – одна из главных загадок зрительного восприятия, о которой мы подробнее поговорим в последних главах.
До 2000-х гг. сетчатка считалась простой нервной системой, состоящей всего из нескольких основных типов клеток. Открытие 29 видов амакриновых клеток и 13 видов биполярных клеток шокировало ученых. На самом деле поначалу эта идея наткнулась на серьезное неприятие. «Да вы, анатомы, просто страдаете маниакальной одержимостью расчленять все на части, – обвиняли нас критики. – Вы считаете, что каждый новый отросток дает вам новый тип клеток». Но доказательства были неопровержимы: особая анатомическая структура клетки, как правило, дополнялась ее особой биохимией и физиологией, а это, в свою очередь, означало, что данная клетка играла в сетчатке особую, отличную от других роль. Другими словами, разные по форме клетки всегда выполняли разные функции.
Прежде существовали и другие намеки на то, что остальная часть нервной системы столь же сложна, но все они по большому счету игнорировались. После публикации моей лабораторией статьи об обнаружении 29 типов амакриновых клеток один авторитетный нейробиолог подсчитал, что в коре головного мозга может существовать около тысячи различных типов нейронов – что намного превышало любые предыдущие оценки[19]. В конце концов ученые осознали, что сетчатка далеко не так проста, как считалось прежде, а остальная нервная система и вовсе умопомрачительно сложна.
На рисунке ниже показаны многие типы клеток, присутствующие в типичной сетчатке млекопитающих. Почему многие, а не все? Потому что уже после создания этого рисунка были открыты новые типы. Тем не менее этот рисунок хорошо иллюстрирует всю сложность нервной системы. Обнаружение такого количества разнообразных типов нейронов кардинально изменило наши представления о том, как функционирует сетчатка глаза: если раньше мы считали, что для понимания работы сетчатки нам достаточно исследовать несколько основных ее составляющих и их комбинаций, то теперь мы встали перед необходимостью изучить около сотни различных микросхем.

Лаконичная максима Стива Куффлера оказалась как нельзя более верной: изучая конкретное, мы действительно узнали кое-что важное об общем – то, что наша нервная система намного разнообразнее и, следовательно, гораздо сложнее в численном плане, чем предполагалось раньше. И узнали мы это, считая нейроны сетчатки.
Часть II
В дебрях мозга
Давайте на минутку остановимся и спросим: что дала нам эта многотрудная работа? Прибыль такова: мы выявили фундаментальный принцип обработки зрительной информации, состоящий в том, что визуальное изображение раскладывается сетчаткой примерно на 30 параллельных потоков сигналов, каждый из которых сообщает мозгу о конкретном аспекте видимого мира. Другими словами, мы воспринимаем видимый мир как совокупность 30 различных параметров, из которых в настоящее время нам известны лишь некоторые: края (контуры), освещенность, движение и цвет. Такой набор из 30 закодированных сигналов передается в мозг о каждой точке видимой картины. Кроме того, вскоре мы поговорим про замечательный набор простых детекторов на входе зрительного проводящего пути в кору мозга. Например, клетки первичной зрительной коры реагируют на такие точные стимулы, как края строго определенной ориентации.
Но насколько все эти знания продвигают нас к нашей конечной цели – ответу на вопрос, как мы узнаем в толпе знакомое лицо, которое может отображаться на сетчатке в сотнях тысяч различных вариантов?
Начиная с первичной зрительной коры, расположенной в задней части мозга, перед нами, как перед древними мореплавателями, простирается таинственный мозговой ландшафт с несколькими известными нам островами, которые мы в какой-то мере исследовали и нанесли на карту, и огромными неизведанными территориями – белыми пятнами на карте, представленной на странице 129, – которые нам еще только предстоит изучить. Но каким образом мы узнали о зрительных областях мозга то немногое, что мы знаем? В основном благодаря экспериментам: записывая электрические сигналы нейронов посредством микроэлектродов или сканируя активность нейронов с использованием технологий визуализации. Так, экспериментальным путем было обнаружено, что в мозге существуют конкретные области, отвечающие за распознавание образов и, в частности, за распознавание видимых объектов, таких как лица (дальше мы поговорим об этом более подробно). Но на сегодняшний день наши знания напоминают отдельные острова на карте – разрозненные факты, которые только-только начинают соединяться не вполне еще четкой сюжетной линией.
Вторая часть книги будет посвящена этой начавшей формироваться сюжетной линии, которую сегодня развивают некоторые ведущие ученые в области нейробиологии и компьютерных наук. Такой подход к пониманию зрительной системы отличается от традиционного, постулирующего иерархию все более специализированных микросхем и не рассматривающего вопрос о том, что именно они собой представляют. Это одна из первых серьезных попыток связать имеющиеся данные в единое целое.
6 | Зрительные сигналы поступают в мозг
Но, чтоб сыграть героя, надо
В грудь его вонзить кинжал по рукоять
И мозг открыто распластать,
Собрать всю яркость красок.
УОЛЛЕС СТИВЕНС
Итак, мы составили примерное представление о выходе из сетчатки: о том, какие зрительные сигналы она передает в мозг. Но что происходит дальше? Можем ли мы картировать маршрут каждого из 30 типов сигналов к их конкретным местам назначения в головном мозге? Оказывается, да, можем – хотя и не для всех. На сегодняшний день мы сумели узнать, куда ведут многие из этих сигнальных путей и что именно происходит в некоторых пунктах назначения. В этой главе я расскажу о нескольких областях обработки зрительной информации в мозге и более подробно остановлюсь на зрительной коре, ключу к загадке распознавания объектов.
ПЕРВЫЕ СТАНЦИИ
Выходные сигналы с сетчатки по аксонам ганглионарных клеток направляются в два основных места назначения в мозге[20]. Одно из них – латеральное коленчатое тело (ЛКТ). Второе – верхнее двухолмие. Ранние анатомы дали этой структуре такое название, потому что она представляет собой два выступа («холмика») на задней поверхности среднего мозга. Как вы могли догадаться, если есть верхнее двухолмие, должно быть и нижнее – эти два нижних выступа ответственны за слух.
Насколько нам удалось выяснить, верхнее двухолмие отвечает в основном за ориентацию зрения. Получая потоки сигналов с сетчатки, эта структура заставляет нас обратить внимание на конкретное место в видимом мире, откуда поступили заинтересовавшие ее сигналы. Если стимулировать электричеством какую-либо точку в одном из холмиков, животное поворачивает голову и глазные яблоки в направлении определенной точки в поле зрения. Повреждение верхнего двухолмия приводит к тому, что животное игнорирует часть поля зрения: происходящее в этой зоне никогда больше не привлекает его внимания.
К сожалению, у нас нет возможности узнать о субъективном зрительном опыте пациентов с поврежденным верхним двухолмием. Поскольку эта структура расположена всего примерно в одном сантиметре над мозговыми центрами, критически важными для сознательной жизни, практически не бывает случаев, чтобы черепно-мозговая травма затронула только верхнее двухолмие и оставила невредимыми нижележащие структуры. Почти всегда повреждаются и эти важнейшие области, а в таких случаях игнорирование части поля зрения – наименьшая из проблем, с которыми сталкивается человек.
Верхнее двухолмие содержит массу интересных нейронных схем со множеством интернейронов и множеством входящих и исходящих нервных путей, связывающих его с другими областями мозга. На самом деле двухолмие имеет слоистую структуру, причем некоторые его слои получают сигналы от слуховой, а не от зрительной системы. Эти слои также отвечают за ориентацию зрения – но они направляют ваш взгляд на источник звука, а не на визуальный стимул. Даже если вы услышите звук с закрытыми глазами, верхние двухолмия все равно заставят вас посмотреть в его сторону. Дело в том, что в природе зрительные и слуховые сигналы часто исходят из одного источника: например, хлопанье крыльев или резкий крик могут свидетельствовать о приближающемся птеродактиле. Синергия зрительного и слухового сигналов позволяет максимально быстро и точно определить местонахождение опасного хищника.
Латеральное коленчатое тело – вторая важная цель зрительного нерва. (Оно называется коленчатым из-за присущего ему небольшого симпатичного изгиба.) ЛКТ представляет собой так называемое ядро – скопление нейронов, которые получают входные сигналы по аксонам зрительного нерва и в свою очередь протягивают множество своих аксонов в зрительную кору. Латеральное коленчатое тело – основная промежуточная станция на пути к зрительной коре. Повреждение этой структуры или нервного тракта между ней и зрительной корой приводит к слепоте в какой-либо части поля зрения. Нервный тракт от сетчатки к латеральному коленчатому телу и далее к зрительной коре – главный проводящий путь сознательного зрения.

Как реагируют нейроны ЛКТ на зрительные сигналы, поступающие к ним по аксонам ганглионарных клеток сетчатки? В ретроспективе ответ кажется очевидным – они должны реагировать так же, как ганглионарные клетки. И действительно, регистрация активности нейронов ЛКТ показала, что они также подразделяются на четыре основных класса: с транзиторными on– или off-ответами и с устойчивыми on– или off-ответами плюс несколько «умных» типов визуальных анализаторов. Все эти сигналы передаются непосредственно в зрительную кору.
Но, как знает каждый студент-нейробиолог, латеральное коленчатое тело – не просто ретрансляционная станция. Со стороны природы было бы бессмысленной глупостью вставлять между сетчаткой и корой целое ядро, которое не имеет никаких других функций, кроме простой передачи сигнала. Анатомические исследования показали кое-что любопытное (и пока необъяснимое): зрительный нерв из сетчатки не является самым большим входом для латерального коленчатого тела. Удивительно, но гораздо больше аксонов – 80 % от общего количества входящих аксонов – идет в коленчатое тело из зрительной коры. Хотя был выдвинут ряд гипотез, никто точно не знает, для чего нужна эта мощнейшая цепь обратной связи. Да, в нейробиологии бывает и такое.
Что же делает латеральное коленчатое тело? Две талантливые исследовательские команды регистрировали активность ганглионарных клеток сетчатки и одновременно их конкретных клеток-мишеней в ЛКТ. Это сложнейший трюк, уверяю вас! Они обнаружили, что у кошек и обезьян эти нейроны-мишени в значительной степени копируют поведение соответствующих нейронов сетчатки. (У мышей происходит почти то же самое, но с подмножеством клеток, получающих очень разные входные сигналы[21].)
В процессе этого ЛКТ усиливает выделение контуров объектов: оно еще больше подчеркивает точки перехода, которые сетчатка сочла необходимым подчеркнуть. Это делается локальными интернейронами, которые встроены в идущий из сетчатки зрительный тракт, по всей видимости, специально с этой целью. На самом деле функция выделения контуров в ЛКТ настолько сильна, что некоторые его нейроны возбуждаются только присутствием краев и почти не реагируют на большие и расплывчатые, то есть не содержащие четких краев, зрительные стимулы.
Еще одно любопытное свойство латерального коленчатого тела состоит в том, что текущий через него поток сигналов может усиливаться или ослабляться под влиянием внешних событий, особенно тех, которые определяют общий уровень возбуждения головного мозга. Когда вы спите, поток информации от сетчатки к зрительной коре резко уменьшается. Это разумно: это все равно что надеть маску для сна, чтобы выспаться в самолете во время ночного полета. Чуть более сложной является способность ЛКТ увеличивать или уменьшать свою пропускную способность в зависимости от избирательного внимания. Когда мы сосредотачиваемся, например, на слушании, ЛКТ помогает нам не отвлекаться на зрительное восприятие. Оно редактирует сообщения, поступающие от сетчатки, и передает в зрительную кору более слабые сигналы о визуальных стимулах.
ОБЫЧНЫЙ ДЕНЬ ИЗ ЖИЗНИ НЕЙРОБИОЛОГА: РЕГИСТРИРУЕМ АКТИВНОСТЬ НЕЙРОНОВ МОЗГА
Разговор о том, что происходит со зрительной информацией дальше – после того как она выходит из латерального коленчатого тела, мы продолжим в следующем разделе. Но сначала я хотел бы рассказать вам, как добываются все те факты, которые я здесь излагаю. Давайте проведем один день в лаборатории, чтобы понять и почувствовать, что же такое нейробиологическая наука на практике.
Конечно, двух одинаковых научных лабораторий не бывает: в каждой свои заведенные порядки, свои конкретные задачи, своя специфика. Некоторые из фантастических новых технологий, о которых я расскажу чуть дальше, диктуют свои особые условия. Но, как бы то ни было, многие исследователи работают именно так, как эти молодые постдоки в типичной лаборатории.
Я посвящаю этому целый раздел моей книги, потому что вы нигде больше об этом не прочитаете. В отличие от редакторов художественной литературы, которые представляются мне благодушными и доброжелательными людьми, редакторы научных журналов, по крайней мере в своей профессиональной деятельности, – это авторитарные тираны, которых ученые абсолютно не интересуют как личности и которые позволяют нам пользоваться свободой творчества только тогда, когда мы пишем собственные имена. В своих вотчинах они устанавливают строжайшие стилистические законы, которые призваны обеспечить максимальную точность и компактность передачи научной информации, не оставляя места для субъективности и допуская лишь редкие (и тщательно маркированные) вкрапления собственного мнения. Авторам обычно выделяют не больше десяти страниц, чтобы изложить результаты нескольких лет работы. Понятное дело, здесь не остается места для описания личного опыта и всех тонкостей проведения экспериментов. В результате у обычных людей складывается превратное представление о том, как делается наука и научные открытия. Чтобы восполнить этот пробел, я предлагаю вам поработать один день вместе с нами в настоящей научной лаборатории.
Мы, постдоки, прибываем в лабораторию к 9:00. Наш руководитель всегда является на несколько минут позже. Поздоровавшись с нами, он проходит к своему столу и погружается в работу – читает и пишет, вполглаза наблюдая за тем, что мы делаем. Наш босс – доктор наук, профессор и опытный экспериментатор, благодаря чему он и стал директором лаборатории, – но на начальном этапе всю грязную практическую работу делаем мы, постдоки. Мы зовем его, только если обнаруживаем что-то интересное и необычное… ну, или если у нас возникают проблемы с аппаратурой.
Наша лаборатория состоит из трех комнат. Первая, площадью примерно шесть на шесть метров – общая рабочая зона. В центре стоит хирургический стол с огромным операционным светильником над головой. Вдоль задней стены – длинный лабораторный стол с черной каменной столешницей и врезанной в нее большой раковиной. Над столом висит ряд шкафчиков со стеклянными дверцами, в которых хранятся хирургические инструменты и разные мелкие приспособления, необходимые для работы. У боковой стены стоит книжный шкаф с полками, забитыми научными журналами. Отдельные полки выделены под лабораторные журналы – одинаковые тетради с бледно-зелеными твердыми обложками и красными тканевыми корешками. Это бесценные документы – в них зафиксирована вся работа лаборатории, ее история. Новые журналы обычно стоят справа.
Регистрирующая аппаратура находится в небольшой смежной комнате. Там стоят три высоких вертикальных стеллажа, заполненных различными электронными приборами. Мы включаем их с утра, как только приходим в лабораторию, потому что оборудованию нужно время, чтобы нагреться. Закончив со всеми необходимыми приготовлениями, мы помещаем подопытное животное, находящееся под глубоким наркозом, в фиксаторы на операционном столе и приступаем к работе.
Сегодня у нас простая задача: узнать, как латеральное коленчатое тело реагирует на сигналы сетчатки. Нейроны латерального коленчатого тела просто повторяют сигналы нейронов сетчатки? Или же они модифицируют сигнал, прежде чем передать его в зрительную кору? Мы не стремимся подтвердить какую-то конкретную гипотезу. Конечно, у каждого из нас могут быть свои личные идеи на этот счет, но они никак не влияют на объективность эксперимента. Мы просто наблюдаем.
Электрическую активность нейронов ЛКТ мы отслеживаем с помощью микроэлектродов. Современные методы позволяют это делать абсолютно безболезненно. Хотя животное находится в глубоком бессознательном состоянии, его зрительная система все равно реагирует на раздражители – сенсорные нейроны возбуждаются, генерируя серии импульсов, когда соответствующий стимул превышает пороговое значение.
Импульс, или спайк, – это электрическое событие, которое можно обнаружить с помощью высокочувствительного электрода, помещенного рядом с клеткой. Чтобы зарегистрировать сигнал одиночной клетки, а не всех ее соседей, электрод должен быть совсем крошечным. Тело нейрональной клетки, где лучше всего записывать сигнал, составляет в диаметре от 5 до 30 мкм. Поскольку в мозговом ядре, таком как латеральное коленчатое тело, нейроны упакованы очень плотно, вам нужно как можно теснее прижать микроэлектрод к конкретному нейрону, чтобы выделить его сигнал на фоне сигналов окружающих клеток.
Итак, мы берем микроэлектрод. (Сегодня микроэлектроды покупаются в основном у коммерческих поставщиков. Но еще до недавнего времени мы изготавливали их сами: для этого бралась тонкая металлическая проволока, которая истончалась посредством травления в электролите и покрывалась изоляционным материалом – пластиком, лаком или стеклом – так, чтобы остался лишь крошечный металлический кончик длиной один-два микрометра. Вся эта операция производилась под микроскопом, чтобы обеспечить высокую точность.) Выводной конец микроэлектрода мы присоединяем к усилителю, а сам микроэлектрод помещаем в специальный микроманипулятор.
Благодаря тончайшему диаметру этот длинный провод можно ввести в мозг, не нанося больших повреждений. Нейроны головного мозга нечувствительны к боли. (Когда у вас болит голова, эти ощущения исходят от окружающих тканей и кровеносных сосудов, а не от нейронов.) Сегодня нейрохирурги лечат некоторые болезни у людей методом так называемой глубокой стимуляции мозга, когда в мозг пациента вживляются крошечные нейростимуляторы. При проведении этой процедуры пациенты обычно находятся в сознании и сообщают врачу о своих субъективных ощущениях, и они никогда не говорят, что испытывают боль, когда тонкая проволока движется внутри их мозга. Это звучит ужасно, но на практике оказывается на удивление безобидной процедурой; на сегодняшний день проведены уже тысячи таких операций, главным образом людям с болезнью Паркинсона, которым такая нейростимуляция помогает смягчить двигательные нарушения.
Но вернемся к нашему эксперименту. Прежде чем подвести микроэлектрод к конкретному нейрону, нам нужно ввести его в латеральное коленчатое тело. ЛКТ невозможно увидеть: оно находится глубоко внутри мозга и закрыто мозговыми полушариями. Поэтому для его локализации мы используем аппарат, называемый стереотаксической рамой. Этот аппарат позволяет определить точное положение мозга относительно находящихся на черепе ориентиров. Опираясь на специальные стереотаксические атласы, где в трехмерной системе координат указаны координаты различных структур мозга, мы пытаемся найти микроэлектродом ЛКТ. Но эти атласы несовершенны, к тому же существует (как у животных, так и у людей) значительная вариабельность соответствий между формой черепа и геометрией мозга, поэтому попасть в нужную нам структуру не так просто.
Обычно требуется несколько попыток. Мы начинаем вводить микроэлектрод в верхней части мозга в точке с координатами X и Y, расположенной вертикально над ЛКТ. Очень медленно, поворачивая микрометрический винт на манипуляторе, мы опускаем электрод все глубже, пока не достигаем указанных в атласе координат ЛКТ. Как мы узнаем, что попали в нужное место? В ответ на световую стимуляцию глаза наш электрод начинает улавливать электрические разряды. Мы отслеживаем их двумя способами. Во-первых, они выводятся на осциллограф – прибор с экраном как у старого телевизора. Это дает нам визуальное отображение активности нейрона – горизонтальную линию с вертикальными зубцами разной частоты. Поскольку изображение на экране лучше видно, когда на экран не падает яркий свет, в комнате полутьма.
Во-вторых, сигналы усиливаются и выводятся на стандартную аудиоколонку (через такие обычно слушают музыку). К счастью, сигналы нейронов звучат в диапазоне частот, которые способен воспринимать человеческий слух. В ходе экспериментов мы контролируем свои манипуляции в основном по звуку, а кривую на осциллографе используем как дополнительный способ контроля. Усиленный одиночный разряд нейрона звучит как короткий хлопок. Множество почти одновременных разрядов – указывающих на то, что мы недостаточно близко подвели микроэлектрод к клетке, – издают потрескивающее шипение. В этом случае на осциллографе мы видим сплошной частокол из небольших вертикальных зубцов. На научном языке это называется неразрешаемой фоновой активностью. Мы же в обиходе говорим «трава» или «шум» – например: «Черт, мы теряем эту клетку в шуме!» В прежние времена нейронограммы записывались на магнитную ленту или фотографировались на пленочный фотоаппарат прямо с экрана осциллографа; сейчас все происходит в цифровом формате.
Крайне редко нам удается сразу же услышать четкий сигнал одной клетки. Обычно мы слышим целый клеточный хор, поскольку все нейроны находятся примерно на одинаковом расстоянии от кончика электрода и ни один из голосов не звучит сильнее других. Изменение общего паттерна нейронной активности сигнализирует нам о том, что наш микроэлектрод достиг латерального коленчатого тела. Чтобы убедиться в этом, мы используем примитивный инструмент – фонарик, работающий от двух батареек. Когда мы быстро проводим лучом света по глазу животного, «трава» на осциллографе становится еще гуще, а колонка издает змеиное шипение: «Ш-ш-ш-ш». Это говорит о том, что мы почти у цели.
Теперь оператор продвигает электрод еще медленнее, аккуратно поворачивая микрометрический винт на манипуляторе. Обычно постдоки работают в паре. Один наблюдает за экраном, чтобы не пропустить момент, когда из «травы» вырастет одиночный шип; другой медленно двигает микроэлектрод, стараясь приблизить его к нейрону. Мы оба внимательно прислушиваемся к исходящему из колонки шипению, ожидая, когда раздастся слабый хлопок. Периодически мы останавливаемся и ждем одну-две минуты: мозговая ткань прилипает к электроду, и эта короткая остановка нужна для того, чтобы ткань сдвинулась вверх на свое место. Еще одна эффективная техника микроперемещения – слегка постучать по столу, на котором лежит животное, чтобы эта легкая вибрация передалась на электрод. Но часто одиночный сигнал возникает из шума сам: услышав слабый хлопок, оператор с предельной осторожностью старается приблизить кончик микроэлектрода вплотную к клетке. Если двигать электрод слишком быстро, можно повредить клеточную мембрану – и убить клетку. Когда такое случается, клетка издает агональный крик из высокочастотной пачки спайков, которая быстро угасает подобно воплю жертвы, падающей с небоскреба в плохом фильме: «Ааааииииииииееее!!!» Но, если мы все делаем правильно, в качестве вознаграждения мы слышим песню одиночного нейрона: ровную барабанную дробь спайков с резкими высокочастотными всплесками, когда мы светим в глаза животного лучом фонарика.
Уже середина дня. После того как мы изолировали одиночный нейрон, наша задача меняется. Теперь нам нужно узнать: что именно этот конкретный нейрон сообщает мозгу о видимом мире? Эксперимент превращается в игру-угадайку. Мы помещаем перед глазами животного светопрозрачный пластиковый экран площадью квадратный метр, на который наклеена тонкая чертежная калька. Активность клетки мы отслеживаем в основном на слух. В полной темноте нейрон все равно генерирует спонтанные разряды в своем уникальном темпе. Зарегистрировав его самопроизвольную активность, мы приступаем к решению нашей задачи – определить, на какие стимулы, паттерны и движения реагирует эта клетка. Мы берем фонарик поменьше – фонарик-авторучку, дающий световое пятно диаметром чуть больше сантиметра. Быстро двигая лучом по экрану (и, следовательно, по сетчатке), мы прислушиваемся к интенсивности импульсов. Приблизительно локализовав чувствительную область – рецептивное поле нейрона, мы берем фонарик с почти точечным пятном света диаметром около двух миллиметров. Снова перемещая это точечное пятно по экрану, мы более точно локализуем рецептивное поле – и аккуратно очерчиваем его границы карандашом на кальке. Эту кальку мы затем вклеиваем в лабораторный журнал, где составляем отчет об эксперименте.
Каждой идентифицированной таким образом клетке мы присваиваем имя с указанием даты эксперимента и порядкового номера, под которым та была исследована в этот день. Но мы пока выяснили только то, за какую область в общем поле обзора отвечает эта конкретная клетка. Следующим шагом мы тестируем ее на избирательность к направлению: мы перемещаем пятно света в пределах ее рецептивного поля, меняя направление движения, скорость, а также размер светового пятна. Если клетка, кажется, предпочитает какое-то одно направление, мы тщательно уточняем, какое именно, и отмечаем его стрелкой на кальке. Если же у нее нет никаких предпочтений такого рода, мы делаем вывод, что это, вероятно, классическая ганглионарная клетка, и переходим к идентификации ее основного типа: on– или off-клетка, с транзиторным или устойчивым ответом. Наконец, мы исследуем механизм латерального торможения. Для этого мы используем два точечных пятна света, одно в центре рецептивного поля, другое непосредственно за его пределами. Сначала мы регистрируем ответ клетки только на световое пятно в центре, затем – только за пределами рецептивного поля и наконец – на два пятна сразу (точность синхронизации обеспечивается электроникой). Практически всегда двойная стимуляция дает более слабый ответ, чем стимуляция только центра, – результат действия латерального торможения.
Иногда нам попадаются крепкие орешки. Они не реагируют на световой стимул ни в одной части поля обзора, и никакие наши ухищрения – изменения направления движения, скорости, размера светового пятна – не заставляют их разразиться высокочастотными пачками спайков. Когда нам не удается вызвать выраженный ответ, есть два варианта: либо клетка повреждена (возможно, мы задели ее оболочку микроэлектродом), либо мы не знаем, на какой аспект видимого мира она реагирует. Если никакие усилия не дают результата, нам не остается ничего другого, кроме как признать поражение и зарегистрировать клетку с удручающей пометкой «неклассифицированная».
О каждой исследованной нами клетке мы делаем короткую запись (от руки, обычной чернильной ручкой) в лабораторном журнале с прошитыми и пронумерованными листами, чтобы ни у кого не возникало соблазна вырвать из него страницу. Если какая-то запись оказалась ошибочной, ее зачеркивают тонкой линией – стирать ее или удалять иным способом не разрешается, чтобы последующие читатели знали, что здесь присутствовала неопределенность.
Наши записи – образец простоты и лаконичности. Вот, например, запись от 15 июня 1985 г.: «Клетка 15/06/85-5, круглое рецептивное поле, on-центр, торможение по окружности. Клетка 15/06/85-10, избирательность к направлению, предпочтительное направление 7:00 → 1:00. Клетка 15/06/85-14, слабая реакция на рассеянный свет, более выраженная реакция не обнаружена. Повреждена?» Эти записи вместе с зарисовками рецептивных полей, записями активности нейронов на магнитных лентах и фотографиях с осциллографов составляют основную базу данных для дальнейших исследований.
Шкаф со священными лабораторными журналами никогда не запирается на замок. Подделать данные экспериментов сложно – да и кому это нужно? Мы не стремимся подтвердить какую-то конкретную теорию, на кону у нас не Нобелевская премия, поэтому в фальсификации результатов нет никакого смысла.
Если все это кажется вам легким делом, вы ошибаетесь. Случается, что аппаратура дает сбой. Иногда приходится тратить массу сил на борьбу с внешними помехами. (Поскольку микроэлектрод – это, по сути, антенна и мы используем очень большое усиление, аппаратура может улавливать 60-герцевую частоту электрического тока в комнатной проводке или даже звуковую дорожку какого-нибудь телеканала. Чтобы избавиться от этих внешних сигналов, приходится перемещать провода или устанавливать в нужном месте защитный экран.) А бывают дни, когда по неясной причине – возможно, вследствие комбинации множества наших собственных мелких ошибок – нам не удается изолировать ни одну клетку. Из-за всех этих и других сложностей за день нам удается успешно исследовать в среднем не больше полудюжины клеток. Обычно эксперимент длится с 9:00 до 18:00. Если дела идут особенно хорошо, мы продолжаем работать допоздна, чтобы исследовать как можно больше клеток. Чтобы составить достаточно полное представление об участке зрительного проводящего пути, нам требуется выборка из нескольких сотен клеток, поэтому весь проект занимает много месяцев. Многие лаборатории проделали эту трудоемкую работу, и она позволила нам сформировать основы понимания того, как работает зрительный анализатор. Путь к пониманию зрения оказался очень медленным и долгим.
ДЕТЕКТОРЫ ОРИЕНТАЦИИ КРАЕВ
Мишень нейронов ЛКТ – первичная зрительная кора – делает кое-что удивительное для системы, которая по-прежнему состоит всего из нескольких видов обрабатывающих элементов: она добавляет новый вид обработки, который вызвал огромное волнение, когда был открыт в 1960-х гг. Я до сих пор помню момент, когда впервые услышал об этом открытии, болтая с коллегами перед семинаром в Гарварде. Тогда мы считали, что в этой области довольно простые цепи нейронных ответов. То, что нейрон, находящийся так близко к периферии зрительного анализатора, может определять ориентацию линии, стало для нас шоком. Это открытие легло в основу модели распознавания визуальных объектов, которая доминировала на протяжении нескольких десятилетий.
Наши коллеги Дэвид Хьюбел и Торстен Визель обнаружили в первичной зрительной коре тип клеток, который не давал выраженной реакции на любую стимуляцию световым пятном. Однако они очень хорошо реагировали на длинную прямую полосу света – линию или край объекта. Что еще более удивительно, эта линия или край должны были иметь определенную ориентацию. Хьюбел и Визель назвали эти нейроны «простыми» клетками (очевидно, потому что они были проще, чем другой тип клеток, о котором пойдет речь дальше). На рисунке на следующей странице представлена нарисованная ими схема реакции простой клетки на светлую линию или край.
Каждый вертикальный штрих на горизонтальной линии обозначает одиночный спайк. Рецептивное поле у простой клетки имеет узкую вытянутую форму. Когда стимул полностью совмещается с рецептивным полем, клетка дает интенсивный ответ. Если стимул находится чуть в стороне, ответ бывает слабым или вовсе отсутствует. Важно отметить, что, если стимул располагается под углом к рецептивному полю, клетка молчит. Другими словами, клетка реагирует только на линии и края строго определенной ориентации, которые находятся в строго определенном месте поля обзора.

Клетка слабо реагирует на равномерное освещение, поскольку возбуждающий и тормозной входные сигналы уравновешивают друг друга. То же самое происходит, когда ее рецептивное поле пересекает линия или край с другой ориентацией: возбуждение и торможение взаимно нейтрализуются. Клетка возбуждается, только когда в ее рецептивное поле попадает линия с точно такой же ориентацией. Короче говоря, это избирательная к ориентации клетка.
Чем полезна избирательность в отношении ориентации? Ответ все тот же: она экономит ресурсы благодаря уменьшению объема информации, передаваемой на следующие этапы зрительной обработки (в данном случае в высшие корковые отделы). Важно отметить, что это делается с сохранением значимой информации, необходимой для идентификации объекта. Самые важные визуальные стимулы – это объекты, а объекты определяются своими краями. Во многих случаях мозгу достаточно знать об ориентации краев – и ни о чем больше, – чтобы догадаться, какой перед ним объект.
На правом рисунке на следующей странице показаны только края – контуры собаки, – которые регистрируются простыми клетками первичной коры. Несмотря на то, что в контурном рисунке частично теряется богатство исходного изображения, собака остается легкоузнаваемой. Эти корковые нейроны делают огромный шаг вперед в процессе извлечения признаков, начатом в сетчатке. Данный конкретный признак, ориентированный край, создает упрощенное схематическое изображение объекта, передача которого в разы более экономичный процесс, чем передача полного изображения. Это можно сравнить с векторным и растровым способом представления изображений в компьютерной графике. Изображения в векторном формате имеют гораздо меньший объем и передаются гораздо быстрее, чем растровые, в которых записывается и передается каждый пиксель. Растровые изображения – самый полный, но и очень неэффективный способ передачи графической информации.

Второй тип клеток, который Хьюбел и Визель назвали «сложными» клетками, также реагировал на линии и края определенной ориентации, но был менее требовательным к их местоположению. Эти клетки возбуждались, когда край имел правильную ориентацию, но был привязан к более широкой зоне. На рисунке, как и в предыдущем примере, вертикальные штрихи обозначают отдельные спайки, генерируемые клеткой, когда в ее рецептивное поле попадает линия с правильным наклоном. Когда же линия имеет другой угол наклона, клетка молчит.

Итак, обобщим: простая клетка возбуждается светлым (или темным) краем определенной ориентации, который находится в конкретном месте поля обзора. Сложная клетка также чувствительна к краю определенной ориентации, но с некоторой степенью свободы: она возбуждается, если такой край появляется где угодно в пределах довольно обширного рецептивного поля, а не только в узкой области.
Вышесказанное важно, потому что эти клетки, по сути, занимаются вычленением такого абстрактного признака, как линейность, до некоторой степени не привязанного к конкретному визуальному стимулу. Хотя их рецептивные поля все равно ограничены, эти клетки выявляют линейность в относительно обширной области, а не в конкретном месте. Это возвращает нас к проблеме, упомянутой в начале книги: к нашей способности распознавать букву А независимо от того, где ее изображение падает на центральную область нашей сетчатки. В 1960-х гг. был разработан последовательный иерархический метод распознавания более сложных объектов, основанный на переходе от обычных неориентационных клеток к простым и далее к сложным избирательным в отношении ориентации клеткам. Эта модель работала не очень хорошо, однако сам механизм, отличающий сложные клетки от простых, сыграл ключевую роль в изобретении важного типа компьютерного зрения (подробнее обо всем этом мы поговорим чуть позже).
Дальше наш путь лежит в почти неизведанные края – в кору головного мозга. Наше понимание коры, говоря откровенно, находится на детсадовском уровне. Пока у нас есть лишь отдельные островки знаний – знаний об отдельных корковых областях, функции которых нам хотя бы приблизительно известны. К счастью, постепенно эти островки начинают соединяться в единый ландшафт – очень грубую, но все же целостную картину того, как организована система зрительного восприятия в головном мозге.
7 | Что дальше: лоскутное одеяло зрительной коры
Есть известные неизвестные вещи – те, о которых мы знаем, что мы их не знаем. Но есть также неведомые неизвестные вещи – такие, о которых мы не знаем, что мы их не знаем… Последние, как правило, представляют наибольшую трудность.
ДОНАЛЬД РАМСФЕЛЬД
Неврологи и нейробиологи считают, что кора головного мозга является средоточием всех тех способностей, которые делают нас людьми, – таких как мышление, речь и чувства. Неудивительно, что зрительная кора в последние десятилетия так интересует исследователей, занимающихся изучением зрения. Важный прорыв был сделан в конце 1990-х гг. вследствие усовершенствования безболезненных способов регистрации нейрональной активности у обезьян, которые находились в сознании и выполняли определенные действия. Это дало нам возможность соотнести конкретные визуальные стимулы с активностью нейронов в конкретных зрительных областях головного мозга.
До сих пор, говоря о «коре», я имел в виду первичную зрительную кору – основную мишень аксонов, выходящих из латерального коленчатого тела. Первичная зрительная кора, сокращенно обозначаемая V1 (от англ. visual – «зрительная»), – это участок коры, расположенный в затылочной части мозга и занимающий примерно 15 % общей площади поверхности коры. Как вы могли догадаться по термину «первичный», в коре имеются и другие зрительные зоны, обозначаемые V2, V3, V4 и т. д. Кроме того, существует множество других областей, которые также реагируют на визуальные стимулы, но не являются чисто зрительными или же вовсе выполняют не связанные со зрением функции и поэтому формально не относятся к зрительной системе.
Итак, оказалось, что зрительные области в головном мозге похожи на лоскутное одеяло. Разные области по-разному реагируют на визуальные объекты и соединены между собой малопонятной нам схемой связей. Поскольку основной массив исследований был сосредоточен на обезьянах и их зрительная кора на сегодняшний день изучена лучше всего, далее я буду говорить в основном об обезьянах, предварительно напомнив вам о том, что, судя по анатомическим и другим параметрам, человеческое зрение не слишком сильно отличается от обезьяньего.
На рисунке на следующей странице показан головной мозг, на котором отмечены отдельные зрительные области. Каждая из этих областей выполняет свою задачу в системе обработки визуальной информации, такую как распознавание объектов, обнаружение движения и т. д.
Первичная зрительная кора (V1) – основной пункт назначения зрительной информации, поступающей из латерального коленчатого тела. Зоны V2, V3 и V4 постепенно уходят все дальше в мозг; их можно рассматривать как более-менее последовательные звенья в цепи обработки зрительной информации.

Помимо них существует множество других зрительных областей, обозначенных на схеме аббревиатурами, некоторые из которых я расшифрую по ходу повествования[22].
Некоторые исследователи считают, что в зрении так или иначе задействовано около 30 % всего объема мозга. Но что же делают все эти вычислительные центры? Эта система так сложна и многообразна, что моему мозгу не хватает когнитивной мощи охватить всю ее сразу. Поэтому я начну с двух наиболее изученных корковых областей: средней височной области (почти в центре рисунка), которая связана с восприятием движения и замечательной группы пятен в нижней височной коре, чьи клетки специализируются на распознании лиц.
СРЕДНЯЯ ВИСОЧНАЯ ОБЛАСТЬ: ПРИРОДНЫЕ ДАТЧИКИ ДВИЖЕНИЯ
Множество умнейших людей корпело над изучением работы нейронов в средней височной коре. Помните мой рассказ о том, как постдоки регистрировали электрическую активность клеток в латеральном коленчатом теле? Здесь происходит все то же самое: вы подводите микроэлектрод вплотную к нейрону, воздействуете на сетчатку различными видами визуальных стимулов и, отслеживая активность нейрона, пытаетесь выяснить, на какой именно стимул он реагирует. Так вот, исследования показали, что нейроны средней височной коры обладают свойствами, которые мы не обнаруживали ранее.
Прежде всего рецептивные поля средневисочных нейронов оказались намного больше, чем у нейронов сетчатки и даже у нейронов первичной зрительной коры. В зависимости от техники измерений их рецептивные поля в четыре – десять раз превышают в размере поля нейронов первичной коры V1. Это означает, что средневисочные нейроны больше нельзя рассматривать как зрительные пиксели; они занимаются более абстрактными вещами.
Ученые выяснили, что эти клетки делают кое-что удивительное. Большинство средневисочных нейронов избирательны в отношении направления: как и нейроны сетчатки, они сообщают остальному мозгу, что что-то движется в определенном направлении. Но из-за большого размера своих рецептивных полей они (в отличие от нейронов сетчатки) не локализуют движущийся объект – тот может находиться где угодно в довольно обширной области видимого мира, чтобы вызвать усиленное возбуждение. Однако эти нейроны обладают другими весьма полезными свойствами.
В пределах своих рецептивных полей средневисочные нейроны относительно неразборчивы. Они реагируют на движение не только одного большого объекта, но и множества маленьких точечных объектов. По причинам, в которые я не буду здесь вдаваться, нейроны сетчатки плохо реагируют на такой тип раздражителя. Можно подразнить средневисочный нейрон, показав ему облако точек, часть которых движется в одном направлении, часть – в обратном (обычно для генерации таких сложных стимулов мы используем специальную компьютерную программу). Так вот, средневисочный нейрон реагирует только в том случае, если решает, что точек, движущихся в предпочитаемом им направлении, больше, чем точек, движущихся в противоположную сторону.
Итак, эта клетка сообщает остальной зрительной системе, что что-то движется в определенном направлении, не уточняя, что именно движется и в какой именно точке пространства. Что еще любопытнее, некоторые средневисочные нейроны реагируют на движение частично невидимых объектов, например таких как барберполы – «цирюльничьи столбы», которыми цирюльники в прошлые века обозначали свои заведения. Барберпол – это вертикальный цилиндр, вращающийся вокруг горизонтальной оси. Но из-за того, что на цилиндр нанесен узор из косых линий, при вращении создается впечатление, будто линии движутся вверх (или вниз, в зависимости от направления вращения). Разумеется, это зрительная иллюзия; на самом деле каждая точка на поверхности цилиндра описывает абсолютно горизонтальный круг вокруг оси вращения. Специализированные лабораторные эксперименты с барберполом показали, что именно средневисочные нейроны обнаруживают это несуществующее движение полос вверх.
Еще одна очень важная функция средневисочных нейронов состоит в том, что многие из них чувствительны к расстоянию до видимого объекта. Мозг вычисляет расстояние посредством сравнения того, куда именно на сетчатку обоих глаз падает изображение объекта. Если объект находится далеко, его изображение падает на обе сетчатки практически в одном и том же месте. Чем ближе находится объект, тем больше это расхождение. Средневисочные нейроны (и нейроны в некоторых других областях) чувствительны к этой разнице во входных сигналах из обоих глаз. При этом они реагируют только на объекты, находящиеся на определенном расстоянии от смотрящего. Более того, избирательность к расстоянию сочетается у них с избирательностью к направлению (о которой мы говорили выше). Другими словами, отдельный средневисочный нейрон может реагировать, например, только на объекты, которые движутся слева направо на расстоянии плюс-минус шесть метров. Будучи всеядными в отношении размера стимула, во всем остальном нейроны этой зрительной зоны чертовски привередливы.
Эти свойства средневисочных нейронов были прямо подтверждены экспериментальным образом. Уильям Ньюсом и его коллеги из Стэнфорда провели следующий эксперимент: с помощью слабых электрических импульсов, подаваемых через микроэлектроды, они стимулировали средневисочные нейроны с определенной избирательностью к направлению у обезьян, которые находились в состоянии бодрствования и демонстрировали выученное поведение (предварительно обезьян обучили наблюдать за реальными движущимися объектами и сообщать о направлении их движения). Исследователи обнаружили, что стимуляция соответствующих средневисочных нейронов улучшала восприятие движения объектов, находившихся в рецептивном поле клетки.
Прежде чем продолжить, я хочу обратить ваше внимание на один ключевой момент: все эти мозговые центры более высокого уровня соединены между собой многочисленными нервными связями и обмениваются друг с другом информацией никак не в линейной последовательности. На рисунке на следующей странице приведена примерная схема основных нервных трактов, участвующих в восприятии движения и центрированных вокруг средневисочной области коры. Это, мягко говоря, похоже на беспорядочную паутину, где все соединено со всем. И роль большинства этих связей неизвестна – они гораздо менее изучены, чем, скажем, проводящий путь из сетчатки к латеральному коленчатому телу и первичной зрительной коре (V1).

Таким образом, в совокупности нейроны средней височной области проводят довольно сложный анализ движений, происходящих в естественном видимом мире, но как именно это делается на уровне отдельных нейронов, во многом остается неясным. Проще говоря, мы знаем некоторые вещи, которые делают отдельные средневисочные нейроны, но не знаем, зачем они это делают – как их конкретный вклад в анализ визуальной сцены способствует формированию ее окончательного целостного восприятия. На этом мы оставим обсуждение средней височной области, поскольку как таковая она не является частью интересующего нас зрительного пути, задействованного в распознавании объектов.
ЗОНЫ РАСПОЗНАВАНИЯ ЛИЦ В КОРЕ
Итак, первая станция обработки зрительной информации в коре – первичная зрительная кора (V1). Далее идут зоны V2, V3 и V4. Грубо говоря, путь информации из зоны V1 в височную кору пролегает через зрительные зоны V2, V3 и V4, однако эти области плохо изучены, и даже самые блестящие ученые умы, несмотря на все свои старания, пока не сумели определить их конкретные функции. (Забегая вперед, скажу, что эти зоны могут быть аналогом скрытых слоев нейронной сети, функции которых зависят от обучения всей сети. Подробнее об этом мы поговорим в главах 10 и 11.) Здесь же я лишь в общих чертах опишу то, что нам известно об этих зонах, чтобы у вас составилось целостное представление о системе обработки зрительной информации в мозге.
Наиболее явное различие между зонами V1 и V2 состоит в том, что нейроны V2 имеют бо́льшие по размеру – а многие из них и более сложные – рецептивные поля, чем нейроны V1. Как известно, V1 содержит больше простых клеток, настроенных на восприятие строго заданных признаков стимулов (таких как линии со строго определенной ориентацией и локализацией в пространстве). Зона V2 содержит больше сложных клеток, чья реакция на ориентированные линии не так жестко привязана к их расположению в пространстве. Однако это различие относительно: в V1 также находится много клеток со сложными рецептивными полями.
Нейроны в зоне V3 обладают разнообразным набором свойств. Почти все клетки показывают избирательность к ориентации, но многие также избирательны в отношении направления и цвета. Раньше зрительная зона V4 считалась центром «цветного зрения», но затем было обнаружено, что здесь также находятся нейроны с избирательностью к ориентации, движению и глубине. Нам пока неизвестно, имеют ли эти участки коры какую-либо функциональную специализацию. Единственное, что мы можем сказать, так это то, что зрительные зоны V1, V2, V3 и V4 представляют собой некую иерархию, где происходит постепенный переход от более простых к более сложным способам обработки визуальной информации.
Далее в височной коре находится кое-что по-настоящему интересное: системы нейронов с замечательной избирательностью к объектам и, в частности, серия участков коры с нейронами, избирательными в отношении лиц. В целом височная доля напоминает шахматную доску из разнообразных функциональных зон, где чувствительные к лицам участки чередуются с зонами, чувствительными к другим визуальным признакам. Впервые избирательные к лицам нейроны были описаны в конце 1970-х гг. Чарльзом Гроссом и его коллегами из Принстонского университета. Они обнаружили эти нейроны в нижней височной коре, которая с высокой степенью избирательности реагировала на определенные объекты, руки и лица.
На тот момент избирательные к лицам клетки показались большинству нейробиологов чересчур специфичными. К тому же в масштабах височной доли таких нейронов было не так уж много, поэтому полученные Гроссом результаты были встречены с весомой долей скептицизма. Когда вы исследуете височную долю с помощью микроэлектрода, как это делал Гросс, вы находитесь во власти случая: вы изучаете тот нейрон, который оказывается под вашим электродом; вы не можете изучить действительно большое количество клеток, потому что регистрация нейронной активности – дело медленное; и, наконец, участки на то и участки (мы называем их «пэтчами» – или «заплатами»), чтобы покрывать лишь небольшую часть поверхности височной коры. Только с появлением технологий сканирования мозга было подтверждено существование этих участков коры, участвующих в распознавании лиц.
Поначалу многие клеточные нейробиологи, включая Чарльза Гросса и меня, пренебрежительно восприняли МРТ-сканирование как инструмент для нейробиологических исследований. В отличие от точных крошечных микроэлектродов, МРТ показывает только большие области мозга, а первые томографы к тому же имели довольно низкое разрешение. Для надежной визуализации требуются особые навыки и соблюдение массы предосторожностей. Поскольку получаемые с помощью МРТ сигналы очень слабы и подвержены разного рода помехам, изображения подвергаются серьезной обработке. Даже небольшие ошибки при обработке могут привести к ложным результатам (немало таких «открытий» популяризовано в научно-популярных статьях по нейробиологии). Но со временем аппараты стали намного лучше, и магнитно-резонансная томография завоевала признание сообщества клеточных нейробиологов благодаря двум неоспоримым преимуществам: во-первых, она полностью неинвазивна; во-вторых, она позволяет одновременно наблюдать активность значительной части мозга, хотя и с гораздо более низким разрешением, чем микроэлектроды.
МРТ дает возможность заглянуть в мозг живых, бодрствующих людей и животных, не причиняя им никакого вреда. Дело в том, что активно работающая часть мозга нуждается в большом количестве энергии, поступление которой обеспечивается за счет усиления кровотока – что и обнаруживает функциональная магнитно-резонансная томография (фМРТ). Функциональная МРТ позволяет экспериментаторам увидеть, какая часть мозга активна в любой момент времени, в частности при выполнении испытуемым различных видов умственной работы. Участникам эксперимента дают какие-либо задания, показывают картинки, предлагают прослушать звуки и т. д. – и с помощью метода фМРТ наблюдают, какие области мозга участвуют в этой конкретной деятельности.
Перелом наступил, когда несколько лабораторий, прежде всего группа Нэнси Кэнвишер из Массачусетского технологического института, зарегистрировали активность человеческого мозга в тот момент, когда люди смотрели на изображения различных объектов. К своему удивлению, они обнаружили, что, когда испытуемые видели человеческое лицо, у них «загоралось» несколько небольших участков в височной доле. Причем эти участки с высокой степенью вероятности повторялись от испытуемого к испытуемому – то есть это было воспроизводимое биологическое событие, а не искажение, вызванное несовершенством аппаратуры. Их «мозаичность» объясняла, почему Гросс и другие ранние исследователи не смогли достоверно продемонстрировать существование участков коры, ответственных за обнаружение лиц: с помощью микроэлектрода очень трудно выявить такие небольшие зоны, разбросанные по обширной поверхности коры.
У людей и обезьян было обнаружено шесть таких участков: они расположены на поверхности височной доли и идут от ее задней части, соседствующей с первичной визуальной корой (слева на этом рисунке), по изогнутой траектории к передней части височной доли.

Точное расположение этих участков немного варьируется, к тому же некоторые из них расположены под поверхностью коры, поэтому на снимках МРТ подобных этому, где показан вид сбоку мозга небольшой обезьяны, обычно не видно сразу все шесть зон. Тем не менее исследования показали, что каждый из шести участков содержит значительную долю клеток, избирательно чувствительных к лицам – будь то человеческие лица, обезьяньи морды, кукольные лица или даже лица мультяшных персонажей.
Затем Дорис Цао, Маргарет Ливингстон и их ученики научились вводить микроэлектроды точно в эти участки. Сложность состояла в том, что эти участки определяются нейронной активностью, а не нейронной структурой, поэтому нет никаких анатомических ориентиров, по которым можно было бы определить их местонахождение. Таким образом, исследователи сначала локализовали эти участки в мозге обезьян с помощью функциональной МРТ (каждый, кто когда-нибудь пытался заставить обезьяну неподвижно лежать в томографе, скажет вам, что это был настоящий научный подвиг), после чего записали электрическую активность нейронов в этих участках с помощью микроэлектродов.
Оказалось, что все чувствительные к лицам участки содержат нейроны с очень большими рецептивными полями. Контролируемые ими области визуального пространства намного больше, чем у нейронов сетчатки, латерального коленчатого тела и первичной зрительной коры (V1). Как только в этой обширной области пространства появляется лицо, нейрон сигнализирует об этом остальной части мозга. Но между этими шестью участками есть важные различия. Нейроны в самых задних участках – наиболее близких к первичной зрительной коре – чувствительны в отношении определенного положения лица. Чтобы они среагировали на знакомое лицо, то должны смотреть в знакомом им направлении: например, чтобы они узнали лицо вашей любимой бабушки, она должна смотреть на ваше левое плечо.
Следующий участок, судя по всему, реагирует не только на знакомое лицо, но и на его зеркальное отражение. Другими словами, здешние нейроны сделали шаг вперед к ключевому принципу распознавания объектов – инвариантности, частично освободившись от привязки к конкретному ракурсу. И, наконец, последний участок, ближайший к лобной доле, содержит нейроны, которые полностью инвариантны к виду: они узнают лицо с любой стороны и в любом виде. Таким образом, ведущая на сегодня гипотеза гласит, что эти шесть участков представляют собой иерархию, в которой нижние уровни более жестко привязаны к регистрируемому сетчаткой изображению, тогда как высшие уровни выполняют более абстрактный анализ визуальной информации.
Шесть участков распознавания лиц функционируют как единая система. Это было продемонстрировано экспериментами, в ходе которых исследователи сначала учили обезьян распознавать конкретные лица, а затем с помощью микроэлектродов стимулировали тот или иной участок. Во-первых, все эти участки оказались непосредственно связанными между собой: стимуляция одного из них вызывала возбуждение всех остальных. Во-вторых, электростимуляция, нарушавшая нормальную активность нейронов в этих участках, ухудшала способность обезьян к узнаванию лиц – что еще раз подтвердило функциональную специализацию участков.
Все мы знаем, как выглядит лицо, но что именно мы имеем в виду, когда говорим, что нейрон «распознает» лица? Как вы уже догадались, такой объект, как лицо, можно разбить на множество отдельных элементов. Один из них – наличие двух глаз. Другой – начинающаяся прямо под ними более-менее вертикальная линия, нос. Под носом находится горизонтальный овал – рот и т. д. Варьируя элементы на изображениях реальных или нарисованных лиц, экспериментаторы наблюдали за усилением или ослаблением ответа специализирующихся на распознавании лиц нейронов. Так, они обнаружили, что эти нейроны слабее всего реагируют на лицо в левом верхнем углу на рисунке ниже и сильнее всего – на лицо в правом нижнем углу, но все они в той или иной степени реагируют на все представленные здесь изображения, даже если на них отсутствуют некоторые элементы лица.

Наблюдаемые паттерны нейрональной активности навели ученых на предположение о том, что эти нейроны измеряют определенные параметры лица и на основе их совокупного анализа делают вывод, является ли объект лицом или нет. Например, Цао и ее коллеги обнаружили чувствительный к лицам нейрон, который сильнее всего реагировал на набор из четырех критериев: соотношение длины и ширины, расстояние между глазами, расположение глаз и размер зрачков. Ни один из этих параметров в отдельности не срабатывал: только совокупность всех четырех могла убедить нейрон в том, что перед ним находится лицо.
Также было установлено, что на каждом участке распознавания лиц разные нейроны настроены на восприятие определенных характеристик лица. Экспериментируя с нарисованными лицами, исследователи обнаружили, что одни клетки наиболее чувствительны к определенному соотношению длины и ширины лица (например, предпочитают длинные тонкие лица в противоположность округлым), другие – к расстоянию между глазами и т. д. Другими словами, нейроны не столько «распознают» лица, сколько измеряют параметры визуального объекта, затем каким-то образом обобщают эту информацию и решают, является ли данный объект лицом или нет[23].
Но как именно нейроны приобретают избирательную чувствительность к лицам, да еще и с такими тончайшими настройками? Я и многие мои коллеги убеждены, что нейроны обучаются этому. Мы исходим из одного критически важного факта: нейронные сети сенсорных систем обладают чрезвычайной пластичностью. И эта тема заслуживает отдельной главы.
8 | Пластичные чувства
Мой умнейший мозг, что мудрей меня,
И хитрей меня.
Как тебе удается бездельничать?
Научи меня, мой господин!
УИЛЬЯМ КАРЛОС УИЛЬЯМС
Издавна было известно, что наши сенсорные системы обладают таким удивительным свойством, как пластичность. У людей, лишенных одного из чувств, происходит компенсаторное усиление всех остальных: например, у слепых людей обостряется слух и осязание. Кроме того, эксперименты по перцептивному обучению (подобные тем, о которых я расскажу в 9-й главе) показали, что с помощью тренировки человек может, например, значительно повысить остроту своего зрения. Конечно, скептики могут возразить, что в этом случае человек просто развивает навык выполнения конкретной задачи и учится лучше концентрировать на ней свое внимание – и, следовательно, ни о каком реальном улучшении работы сенсорной системы речь не идет. Но исследования с использованием современных методологий убедительно доказали, что при этом происходит физическое изменение нейронных сетей в головном мозге.
ПОВРЕЖДЕННЫЕ СЕНСОРНЫЕ СИСТЕМЫ ПЕРЕНАСТРАИВАЮТСЯ
Термин «пластичность мозга» означает способность мозга к реорганизации своих нейрональных схем. Один из классических ранних экспериментов, показавших это, состоял в следующем: исследователи брали слепую от рождения (например, вследствие повреждения обеих сетчаток) крысу. Когда крыса подрастала, ее обучали проходить лабиринт, что животное делало с удовольствием, потому что в конце лабиринта его ожидало лакомство. Затем исследователи слегка повреждали крысе зрительную кору и снова запускали в лабиринт. Теоретически поврежденная зрительная кора не должна была повлиять на способность зверька ориентироваться в пространстве. Но на деле крыса находила путь в лабиринте гораздо хуже, чем раньше, что означало, что зрительная кора у слепых крыс каким-то образом задействована в выполнении этой задачи, хотя и неизвестно, как именно. Эксперимент был осуществлен, в частности, Карлом Лешли из Лаборатории биологии приматов имени Р. Йеркса, которого интересовало физическое местонахождение памяти в мозге. Последующие исследователи указывали на некоторые недостатки его метода, но в целом ученый двигался в правильном направлении[24].
Еще одно наблюдение, на этот раз касавшееся людей, сделали в те же годы врачи-клиницисты. Они сообщали о двух типах приобретенной слепоты. К первой категории относились пациенты, у которых с рождения был перекрыт один глаз, например, катарактой или вследствие редко встречающегося нарушения развития век. Когда анатомическая проблема в конце концов устранялась, глаз все равно оставался слепым или почти слепым. Что-то происходило в течение этого раннего периода окклюзии, что затем не давало глазу и его проводящему пути в центральной нервной системе начать функционировать надлежащим образом.
Ко второй категории относились дети с врожденным косоглазием. Когда такие дети вырастали, один глаз у них обычно брал верх над другим, в результате чего второй глаз переставал работать. В просторечии это называется синдромом ленивого глаза, или спящего глаза, на официальном медицинском языке – амблиопией. На самом деле этот глаз не слепой – специализированные тесты показывают, что его сетчатка вполне функциональна, – просто он не участвует в зрении. (Существуют различные методы лечения этой патологии, наиболее распространенный – попеременное перекрытие глаз в раннем детстве, чтобы ни один глаз не мог взять на себя доминирующую роль и подавлять другой.)
Исследователи Дэвид Хьюбел и Торстен Визель, первооткрыватели простых и сложных нейронов в зрительной коре, провели аналогичные эксперименты на животных и описали нейрональную основу ленивого глаза: оказалось, что в ранний период жизни синапсы, которые соединяют идущий от сетчатки зрительный проводящий путь с центральной нервной системой, очень пластичны. Если корковые нейроны получают основную зрительную информацию от одного глаза и почти не получают ее от другого, аксоны первого глаза захватывают все синаптическое пространство корковых нейронов, из-за чего второй глаз, хотя и остается функциональным, лишается возможности передавать сигналы мозгу.
Кроме того, Хьюбел и Визель обнаружили, что проблема косоглазия гораздо сложнее, чем считалось раньше. В норме изображения с обоих глаз почти идеально совпадают топографически: один и тот же участок визуальной сцены стимулирует одну и ту же группу корковых нейронов. Когда же исследователи вызвали у животных искусственное косоглазие (заставив животных с рождения носить на одном глазу призму, отклонявшую падающее на сетчатку изображение) и затем исследовали их мозг, оказалось, что сигналы из двух глаз возбуждали разные группы нейронов-мишеней, то есть не создавали единую кортикальную карту. Другими словами, при косоглазии зрение в буквальном смысле слова становится бинокулярным: вместо одного изображения мозг видит два разных. А поскольку эти два изображения конфликтуют друг с другом, мозгу приходится выбирать, какому из них отдать предпочтение. В итоге синаптические связи с одним из двух глаз подавляются – сначала временно, а затем и навсегда, делая этот глаз функционально слепым.
Чарльз Гилберт из Рокфеллеровского университета провел изобретательный эксперимент, показавший, как происходит реорганизация корковых ответов другого рода. В норме в зрительной коре существует своего рода «карта» сетчатки. Конечно, она немного искажена из-за неровной поверхности коры, однако исследования показали, что соседние точки на сетчатке проецируют свои ответы на соответствующие соседние точки в зрительной коре, создавая на ней упорядоченную карту поля обзора. Эксперимент заключался в следующем: с помощью лазера в сетчатке обезьяны безболезненно проделывалось крошечное отверстие, после чего Гилберт регистрировал активность ее зрительной коры. Поначалу на кортикальной карте появлялся пробел, соответствовавший отверстию в сетчатке. Но спустя какое-то время этот пробел затягивался: соседние области сетчатки образовывали синаптические связи с клетками зрительной коры, которые остались без работы из-за повреждения их источника сигналов.
Это не означает, что в поврежденной области сетчатки было восстановлено зрение. Если сетчатка повреждена, в этом месте навсегда останется слепое пятно, которое ни сетчатка, ни мозг не смогут компенсировать. Происходит другое: освободившиеся корковые нейроны «захватываются» клетками вокруг места поражения сетчатки, что, в свою очередь, позволяет предположить, что эта область начинает функционировать эффективнее, чем раньше. Насколько мне известно, никто пока не протестировал это предположение. Как бы то ни было, гиперинервированная область сетчатки должна быть более устойчивой – и, возможно, менее чувствительной к дальнейшим повреждениям, чем прежде.
Таким способом природа предотвращает простой вычислительных мощностей. Если какая-то область коры перестает получать входные сигналы из своего естественного источника, она бездействует – а это непозволительная роскошь для нашего мозга. Поэтому через какое-то время эта корковая область переключается на обслуживание неповрежденных источников. Аналогичный механизм предположительно используется мозгом для преодоления последствий мелких инсультов. (Невропатологи утверждают, что мы теряем небольшие участки мозговой ткани на протяжении всей жизни.) Представьте, что в коре произошла закупорка мелкого кровеносного сосуда, в результате чего крошечная область мозга, которую он питал, отмерла. Корковая область-мишень, которая раньше обрабатывала входные сигналы из этого навсегда замолчавшего источника, осталась не у дел. Но природа старается по максимуму задействовать драгоценные корковые ресурсы, поэтому она передает их в распоряжение соседних областей.
РЕОРГАНИЗАЦИЯ НОРМАЛЬНЫХ СЕНСОРНЫХ СИСТЕМ
В предыдущем разделе мы говорили о том, как сенсорные системы приспосабливаются к различным видам патологий и повреждений, серьезно нарушающих их нормальное функционирование. Но небольшие реорганизации систем восприятия – естественный процесс, который происходит с каждым из нас.
Когда исследователи получили доступ к технологиям сканирования мозга, мы узнали много интересного. Яркий пример пластичности мозга был обнаружен при исследовании мозговой активности слепых от рождения людей. Так, когда слепые добровольцы читали с помощью пальцев текст, напечатанный шрифтом Брайля, у них активизировались области мозга (такие как первичная зрительная кора), которые у зрячих задействованы в обработке визуальной информации. Слепые на протяжении многих лет интенсивно использовали свое чувство осязания, в результате чего функция обработки тактильной информации каким-то образом была передана незадействованному зрительному центру.
Другой впечатляющий пример был получен в ходе исследования зрячих людей – профессиональных скрипачей. При игре на скрипке человек делает относительно несложные и неточные движения одной рукой, которой водит смычкой по струнам. При этом другой рукой он должен делать точно выверенные мелкие движения, прижимая струны к грифу скрипки в строго определенных местах (иначе ноты будут звучать фальшиво) – причем иногда это необходимо делать с головокружительной скоростью. Другими словами, здесь сочетается высокая скорость и точность движений. Профессиональные скрипачи упражняются по многу часов в день. Как вы уже догадались, это отражается на физической организации связей в их головном мозге – конкретно в той области мозга, которая контролирует движение пальцев. У профессиональных скрипачей эта область расширяется настолько, что даже заменяет функции соседней мозговой ткани. Но это происходит только для одной руки, используемой для нажима на струны. Та же область в другом полушарии, которая контролирует относительно простые движения руки, работающей смычком, не меняется.
(Мне интересно, не происходит ли с другими профессионалами то же самое, что и со скрипачами. Например, не могут ли у профессиональных спортсменов расширяться области мозга, отвечающие за контроль определенных мышц? Или, если вы всю жизнь изучаете мозг, не приводит ли это к тому, что «область изучения мозга» в вашем собственном мозге расширяется настолько, что вытесняет все прочие нейронные сети, например, отвечающие за любовь к опере?)
Ситуация, противоположная интенсивному использованию сенсорной системы, называется сенсорной депривацией. Эксперименты показали, что у кошек, выросших в полной темноте, мозг не развивал способность сочленять монокулярные изображения в единое целое. Чуть более спорные результаты дали эксперименты с выращиванием кошек в условиях, когда единственными доступными для зрительного восприятия ориентированными раздражителями были вертикальные или горизонтальные полосы. Избирательность к ориентации вообще слабо выражена от рождения и развивается в постнатальный период. Взросление, проведенное в «полосчатой» среде, отражается на соотношении избирательных к ориентации нейронов в первичной зрительной коре: у кошек, выросших в окружении вертикальных полос, было обнаружено аномально большое количество нейронов, чувствительных к горизонтальной ориентации; у кошек, видевших только вертикальные полосы, такой же перекос наблюдался в сторону чувствительных к вертикальной ориентации нейронов[25].
В более продвинутом варианте эксперимента кошек в ранний период жизни лишали возможности видеть движение. Для этого экспериментаторы создали среду, где единственным источником освещения были вспышки стробоскопа. Это позволяло кошкам видеть окружающий мир, однако вспышки были слишком короткими, чтобы клетки сетчатки успевали зарегистрировать значимое движение объектов – другими словами, кора этих животных была лишена визуальной информации о движении. Что произошло? Эти животные выросли без избирательных к направлению нейронов в коре мозга.
Наконец, еще одна важная серия экспериментальных манипуляций, осуществленная Майклом Страйкером и Карлом Шатцем (ныне работающими в Калифорнийском университете в Сан-Франциско и Стэнфордском университете соответственно), непосредственно подтвердила роль синаптической пластичности в развитии зрения – конкретно на примере латерального коленчатого тела.
Как известно, одна часть ЛКТ принимает входные сигналы от левого глаза, другая – от правого. Но такая специализация присуща ЛКТ не с самого рождения. У младенцев аксоны, идущие от обоих глаз, широко ветвятся и охватывают обширные области ЛКТ. Другими словами, поначалу в ЛКТ практически нет физического разделения на правый и левый глаз. Такая сегрегация создается за счет паттерна активности аксонов, идущих от разных глаз. Эти аксоны начинают работать еще до рождения – они генерируют отдельные всплески импульсов подобно автомобильному двигателю, работающему на низких оборотах на холостом ходу. Вспышки синхронизированы: импульсы от одного глаза поступают в ЛКТ в один момент времени, импульсы от другого глаза – в другой.
Тем самым создаются условия для важнейшей формы пластичности – прообраза машинного обучения. Как мы подробнее рассмотрим в главе 9, канадский нейробиолог Дональд Хебб предположил, что совместная активизация нейронов усиливает синаптические связи между ними. В нашем случае аксоны, идущие от сетчатки одного глаза, одновременно активируют свои клетки-мишени в ЛКТ, в результате чего синаптические контакты между этими аксонами и клетками ЛКТ усиливаются по сравнению с синаптическими контактами с аксонами другого глаза. Таким образом, широко разветвленные аксоны постепенно сосредотачиваются на определенных мишенях, так что один кластер нейронов ЛКТ становится чувствительным к сигналам от правого глаза, другой кластер – к сигналам от левого глаза. Механизм Хебба – усиление одновременно активируемых синаптических связей – приводит к уточнению изначально беспорядочной карты сетчатки на ЛКТ. Чтобы проверить это предположение, Страйкер и его коллеги с помощью специального препарата блокировали у животных активность зрительного нерва, исходящего от одного глаза, и уточнения карты не происходило.
Все эти результаты указывают на пластичность организации сенсорных систем. Но может ли это помочь человеку, который первые годы жизни прожил без зрения вообще?
МОЖНО ЛИ НАУЧИТЬСЯ ВИДЕТЬ?
Дональд Хебб предположил, что зрение в значительной степени связано с обучением: сложное зрительное восприятие формируется на основе опыта, посредством ассоциаций, поскольку объекты видимого мира представляют собой совокупности отдельных признаков. Хебб считал, что это обучение происходит в раннем возрасте и что в дальнейшем мозг, как давали предполагать некоторые существовавшие на тот момент данные, не способен объединять такие признаки. В основном он был прав: зрение действительно в большой степени зависит от визуального опыта. Но его вывод о том, что подобный опыт приобретается только в раннем возрасте, оказался справедлив лишь отчасти.
Источником данных служат исследования людей, которые были слепыми от рождения и начали видеть в более позднем возрасте. Во время посещения Индии профессор Паван Синха из Массачусетского технологического института обратил внимание на большое число детей с плотной врожденной катарактой – по его оценкам, в сельской местности их насчитывалось около трехсот тысяч. При этой патологии вместо прозрачного хрусталика в глазу формируется мутная фиброзная ткань, которая пропускает только свет, но практически лишает зрения. Соединив замечательным образом филантропию и науку, Синха организовал программу помощи таким детям: из удаленных деревень их доставляли в современную глазную клинику в Нью-Дели, где хирурги заменяли дефектные хрусталики на прозрачные синтетические линзы (в развитых странах мира это стандартная операция, которая проводится многим пожилым людям со старческой катарактой).
Команда Синха проверяла зрение у маленьких пациентов перед операцией, сразу после нее, а также спустя несколько месяцев и несколько лет. У прооперированных детей не сразу появлялось четкое зрение. Поначалу они видели мир как расплывчатое пятно. Но постепенно четкость и детализация их зрительного восприятия улучшались. Многие в конце концов были способны ходить без трости для слепых, ездить на велосипеде по многолюдной улице, узнавать родных и друзей, ходить в школу и заниматься другими видами деятельности, предполагающими способность видеть.
Однако зрение так никогда и не стало идеальным. Даже после многих месяцев тренировки острота зрения у них оставалась ниже нормы. Некоторые могли прочитать газетные заголовки, но не мелкий шрифт. Многие не могли справиться с определенными визуальными задачами, например, такими как разъединение двух наложенных друг на друга изображений, как показано на рисунке. Зрячие с рождения люди видят здесь треугольник, частично наложенный на квадрат. Но большинство людей, которые обрели зрение в более позднем возрасте, воспринимают это как единый сложный объект. (Интересно, что эта проблема легко «излечивается», если одна фигура движется относительно другой. Этот и другие специальные методы тренировки зрения показали свою полезность для восстановления зрительной функции.)

Итак, зрение в значительной степени поддается восстановлению, но некоторые открытия напоминают нам, что пластичность зрительной системы не безгранична. Например, тот факт, что шесть участков распознавания лиц находятся у людей (и у обезьян) практически в одних и тех же местах, свидетельствует о том, что мозг имеет некоторую заданную на генетическом уровне схему внутренней организации. Это подтверждается и реорганизацией мозга у вышеописанных индийских детей. Сразу после удаления катаракты функциональная МРТ показывала дезорганизованную, охватывающую широкие области мозга реакцию на визуальный ввод, в том числе на лица. Но уже спустя короткое время реакция на лица концентрировалась в серии участков, располагающихся в тех же местах, что и у зрячих с рождения людей. Другими словами, мозг выделил под функцию распознавания лиц не любые случайные зоны, а строго определенные места – что свидетельствует о присущей визуальным структурам как минимум некоторой степени предопределенности. Ливингстон назвала такие предопределенные локации «протоструктурами».
В конце 2017 г. Ливингстон с соавторами опубликовала результаты оригинального эксперимента по изучению пластичности сенсорных нейронов. Группа Ливингстон выращивала обезьян с рождения в среде, где они никогда не видели лиц – ни человеческих лиц, ни обезьяньих морд – вообще никаких. Сделать это было не так трудно, как кажется. Экспериментаторы с любовью заботились об обезьянах, но появлялись перед ними только в маске из темного тонированного стекла наподобие маски сварщика, которая закрывала их лица от лба до подбородка.
В остальном обезьяны росли в совершенно нормальном визуальном мире: в клетках было множество разнообразных предметов; они видели туловище, руки и ноги экспериментаторов, бутылочку, из которой их кормили. Они слышали обычные звуки, которые слышат обезьяны в обезьяньих колониях. Их единственной сенсорной депривацией было то, что они никогда не видели лиц. Животные развивались абсолютно нормально и после завершения эксперимента успешно социализировались со своими собратьями и интегрировались в обезьянье общество.
После того как экспериментаторы приучили обезьян неподвижно лежать внутри томографа, они протестировали их реакцию на различные визуальные стимулы. Как вы могли догадаться, при виде лиц у этих животных не активизировались типичные участки распознавания лиц в височной коре. Но примечательно то, что эти участки мозга реагировали на изображения рук. В нормальной социальной среде наиболее важными визуальными объектами для приматов являются лица. Лица сигнализируют о гневе, страхе, враждебности, любви и других эмоциях, то есть несут важную для выживания информацию. По всей видимости, вторым по важности объектом окружающего мира являются руки их сородичей или в данном случае экспериментаторов, растивших и кормивших подопытных обезьян.
Но нейроны, которые переквалифицировались в распознавание рук, сохранили свою пластичность. Примерно через шесть месяцев после того, как обезьянам разрешили видеть лица экспериментаторов и других обезьян, нейроны на участках распознавания лиц постепенно приобрели избирательную чувствительность к лицам. Очевидно, лица несут так много важной информации, что природа сочла необходимым вернуть этим нейронам на обработку вычислительные мощности, временно «захваченные» руками.
Существование специальных участков распознавания лиц также объясняет такое необычное и давно известное расстройство восприятия, как лицевая слепота или прозопагнозия (от греч. prosop – лицо, и agnosia – неузнавание). При этом расстройстве человек обладает абсолютно нормальным зрением, но испытывает трудности с узнаванием лиц. Он распознает лица и отличает их друг от друга, но не может узнавать знакомые лица по памяти.
Есть разные степени прозопагнозии – от легкой формы до почти полной «лицевой слепоты», которая обычно требует медицинского вмешательства. На противоположном полюсе находятся люди с суперспособностью к запоминанию лиц. Сенатор Эдвард Кеннеди, по словам одного из его помощников, мог узнать по памяти 10 000 человек. Лично я подозреваю у себя легкую форму прозопагнозии. Я могу провести отличный вечер в приятной компании, а на следующий день, столкнувшись с кем-то из вчерашних собеседников, подумать: «Мне кажется, я где-то видел этого человека? Или нет?» Поэтому, если я когда-нибудь пройду мимо вас, не поздоровавшись, не вините меня в спесивости – это недоработка моего мозга.
Итак, подведем итог: зоны распознавания лиц расположены в определенных местах головного мозга; предположительно функционируют как единая система и развивают избирательную чувствительность к лицам посредством опыта. Как указывает Ливингстон, эта распределенная, зависимая от опыта система во многих отношениях ведет себя так, как если бы ее клетки были элементами своего рода обучаемой нейронной сети.
9 | Формирование нейронной сети: нейроны соединяются, если вместе возбуждаются
Моей целью было понять… почему удаление у пациента значительного по размеру участка мозговой ткани почти не отражалось на его интеллектуальных и иных способностях, по крайней мере так, чтобы это стало заметно для членов его семьи. Каким образом удаление всей правой половины коры не приводило к потере интеллекта?
ДОНАЛЬД ХЕББ
Теперь мы начнем сводить все воедино – чтобы наконец-то ответить на вопрос, поставленный в начале книги: как мы узнаем своего ребенка? Для этого давайте вернемся в 1960-е гг., когда на стыке нейробиологии и вычислительной биологии родилась захватывающая новая дисциплина, основы которой были заложены более полувека назад канадским нейробиологом Дональдом Хеббом.
Представьте себе мозг, содержащий всего два нейрона. Как и большинство нейронов, они соединены между собой через синапсы. (Оставим пока в стороне общение с внешним миром. Это упрощенный гипотетический мозг.) Оба нейрона время от времени возбуждаются. В данном случае под возбуждением понимается просто генерация нервных импульсов.

Теперь сравним две ситуации. В первом случае два гипотетических нейрона возбуждаются в разное время: когда один нейрон активируется, другой молчит. Каждый срабатывает в ответ на входные сигналы безотносительно к другому. Во втором случае нейроны срабатывают вместе: когда возбуждается один нейрон, возбуждается и другой. В обоих случаях уровень активности обоих нейронов одинаков. Единственная разница в том, что они срабатывают либо одновременно, либо в разное время.
Оказывается, при асинхронном срабатывании ничего особенного не происходит. Но, когда нейроны возбуждаются вместе, происходит кое-что замечательное: синаптическая связь между ними постепенно усиливается, так что в конце концов активизация одного нейрона начинает вызывать активизацию и другого. Студенты запоминают это правило так: «Нейроны соединяются, если вместе возбуждаются».
Такой вот простой кусочек нейробиологии, который я только что объяснил вам полутора сотней слов. Однако этот механизм – усиление синаптических связей при одновременной активизации нейронов – является самой базовой формой памяти, которая лежит в основе всего нашего восприятия, эмоций и действий. Эту и другие необычайно провидческие идеи о нервных сетях Дональд Хебб, один из основоположников современной нейробиологии, изложил в своей эпохальной работе «Организация поведения», опубликованной в 1949 г. Модифицируемая связь между двумя нейронами получила название синапса Хебба. Позже мы подробно поговорим о потомках этого синапса, используемых в современных нейронных сетях: на синапсах Хебба – вернее, на различных его кремниевых версиях – построено большинство современных форм искусственного интеллекта.
ОРИГИНАЛЬНАЯ НЕЙРОННАЯ СЕТЬ ХЕББА
В работе «Организация поведения» Дональд Хебб изложил очень широкое теоретическое ви́дение, охватывающее многие аспекты нейробиологии. Эта теория не только заложила фундамент того, что сегодня известно как машинное обучение, но и обеспечила понимание того, как животные выживают в естественном мире. Однако главным интересом Хебба, почвой, на которой выросла его теория, было восприятие.
Хебб начал с простейших видов восприятия. Возьмем элементарный рисунок – черный контурный квадрат на белом фоне. Мы видим квадрат как целостную фигуру, нарисованную непрерывной линией. Но в действительности наша сетчатка воспринимает черную линию как ряд точек, потому что изображение линии падает на множество отдельных светочувствительных нейронов – палочек и колбочек. Как бы плотно ни располагались друг к другу фоторецепторы и какой бы ровной ни казалась нам линия, сетчатка все равно регистрирует ее изображение как ряд точек.
Но мы, разумеется, этого не видим. Мы видим черный контурный квадрат на белом фоне, потому что наш мозг соединяет все точки в цельные линии. Более того, мы не воспринимаем каждую из четырех линий по отдельности: мы видим квадрат как единый объект. Однако поврежденный мозг может терять такую целостность восприятия. Нейропсихолог Ханс-Лукас Тойбер из Массачусетского технологического института описал случай, когда пациент вместо одного движущегося мотоцикла видел вереницу отдельных мотоциклов. Нормальный мозг, как правило, видит объекты как единое целое, а не как наборы отдельных компонентов. Но на самом деле ничто в видимом мире не говорит нам о том, что является объектом, а что нет; мы видим лишь массив пикселей, из которых мозг должен вычленить целостные объекты. В примере Тойбера мозг должен понять, что перед ним один движущийся мотоцикл, а не цепочка из неподвижных мотоциклов. Этот принцип, называемый завершенностью, подводит нас еще на шаг ближе к пониманию механизма восприятия, описанного Хеббом.
Еще в начале XX в. психологи предположили, что в основе восприятия объектов лежат их гештальты – целостные и обобщенные образы объектов, игнорирующие вариации в конкретных деталях. Чтобы проиллюстрировать это, обычно используется изображение знакомого предмета с недостающим фрагментом. Посмотрите на этот рисунок: вы мгновенно понимаете, что перед вами треугольник, несмотря на то что одна его сторона не дорисована до конца.

Хебб и его ученики первыми описали одно из интригующих проявлений этого феномена, которое они обнаружили с помощью так называемого метода стабилизации изображения на сетчатке. Хотя мы этого не осознаем, наши глаза постоянно совершают очень мелкие высокочастотные колебания, называемые «тремор глаз» (не путайте их с совсем другим явлением – скачкообразным движением глаз, когда мы сканируем визуальную сцену). Эти мелкие движения, которые мы даже не замечаем, играют важную роль в функционировании нашей зрительной системы. Чтобы стабилизировать изображение на сетчатке, Хебб и его команда сконструировали особые контактные линзы, из центра которых выходила тончайшая ножка с еще одной крошечной линзой на конце. Линза фокусировала тестовое изображение на сетчатке и таким образом устраняла эффект тремора глаз.
Большинство нейронов сетчатки и зрительных отделов мозга не очень интересуются стабильными объектами, которые не меняются и не движутся. Они активно отвечают на появление в поле зрения нового объекта, но, если в его изображении не происходит никаких изменений, перестают реагировать. Это полезное свойство, которое позволяет мозгу не тратить драгоценную энергию на то, что не несет новой информации. Его побочный и не очень полезный эффект состоит в том, что, если объект не движется, мозг попросту перестает его замечать. Тремор глаз в состоянии покоя противодействует этому, заставляя изображение постоянно перемещаться туда-сюда по сетчатке (повторю: эти движения слишком малы, чтобы мы их замечали), поэтому нейроны не перестают на него реагировать. Использованные Хеббом специальные контактные линзы устраняли эффект этих мелких движений глаз, при каждом движении сдвигая падающее на сетчатку изображение.
Как вы могли догадаться, при использовании экспериментальной контактной линзы изображение постепенно исчезало, так как в отсутствии тремора не происходило активации новых рецепторов. Но для Хебба самым интригующим результатом было не исчезновение изображения, а то, каким образом оно исчезало. Это не было хаотичным процессом, например, в виде превращения в беспорядочную россыпь точек. Стабилизированное изображение исчезало отдельными значимыми фрагментами. Сплошной квадрат мог исчезнуть сразу, но контурный квадрат сначала лишался одной стороны, затем еще двух, после чего исчезала последняя четвертая сторона.
Хебб предположил, что эти фрагменты кодируются группами одновременно активизирующихся нейронов в головном мозге. Он назвал эти группы, по сути, представлявшие собой базовую версию нейронной сети, ансамблями клеток. Кроме того, Хебб выдвинул гениальную идею относительно того, как образуются клеточные ансамбли, – этот механизм, в существование которого ученый верил, впоследствии был назван синапсом Хебба (чуть дальше мы поговорим о нем очень подробно).
Теория клеточных ансамблей Хебба дала два важных результата. Во-первых, она объясняла, почему восприятие происходит отдельными значимыми фрагментами. Изображение квадрата в мозге активирует четыре ансамбля клеток (по одному для каждой из четырех сторон), соединенных синаптическими связями. При этом вовсе не обязательно, чтобы эти четыре ансамбля физически располагались в мозге также в форме квадрата. (На самом деле эти базовые клеточные ансамбли и их ансамбли-мишени более высокого уровня могут быть распределены по всему мозгу.) Согласно разработанной Хеббом схеме, за восприятие таких простых элементов, как квадрат, отвечают самые базовые клеточные ансамбли, которые, в свою очередь, связаны между собой для восприятия более сложных объектов и даже для формирования мыслей и сознательной памяти. Этот конкретный квадрат представляет собой сенсорную репрезентацию из четырех линий, но через связь с другими клеточными ансамблями становится частью хранящегося в памяти образа квадрата.
В этой книге мы ограничимся рассмотрением простых форм восприятия, однако следует отметить, что клеточные ансамбли Хебба могут быть связаны не только с другими ансамблями того же типа (например, зрительными), но и с клеточными ансамблями, отвечающими за восприятие звуков, запахов или вкуса. Эмоции – еще одна важнейшая паутина таких ансамблей, которые хранят историю каждого человека и могут быть связаны с очень давними воспоминаниями. Понятно, что подобного рода ансамбли могут насчитывать сотни тысяч и даже миллионы взаимосвязанных нейронов, находящихся как близко, так и очень далеко друг от друга. Именно такие обширные паутины связей лежат в основе концептуальных и эмоциональных ассоциаций. Если вы читали Пруста, теперь вы понимаете, почему вкус печенья «Мадлен» вызвал у главного героя нахлынувшую волну воспоминаний о «потерянном времени».
Такая паутиноподобная ассоциативная организация – первое, что многим приходит на ум, когда мы слышим термин «нейронная сеть». И, если смотреть издалека, все множество связей в мозге действительно выглядит именно так. Но в этой паутине есть высокоспецифичные связи, и именно они обеспечивают основу зрительного восприятия.
Второе следствие теории Хебба не имело отношения к восприятию, но было не менее захватывающим. Его идеи объясняли удивительную устойчивость мозга к повреждениям. В 1937 г. монреальский хирург Уайлдер Пенфилд начал применять новую процедуру хирургического вмешательства для удаления очагов судорожной активности в головном мозге у больных эпилепсией. Эти очаги представляют собой участки коры с аномально активными нейронами, которые распространяют возбуждение на нормальные части коры и тем самым вызывают эпилептические припадки. Их удаление во многих случаях позволяет значительно сократить частоту приступов. Но если при резекции аномальных очагов затрагиваются сенсорные или моторные зоны коры, пациент может частично или полностью потерять зрение, слух, способность ходить или выполнять сложные движения какой-либо частью тела. Чтобы не допустить этого, Пенфилд во время операций на открытом мозге стимулировал электричеством различные части коры – при этом пациенты оставались в сознании и описывали свои ощущения, – чтобы точно локализовать специфические сенсорные и моторные области и не повредить их при удалении эпилептического очага.
В конце концов он составил функциональную карту коры мозга, обозначив на ней все идентифицированные сенсорные и моторные зоны. Однако значительная часть поверхности осталась на карте белым пятном. Пенфилд никак не мог определить функции этих областей (они остаются плохо изученными и сегодня). Тогда он обратился за помощью к Дональду Хеббу, чтобы тот попытался выяснить, к каким именно нарушениям приводит резекция поврежденных тканей в этих «молчащих» областях. Другими словами, если удалить конкретный сегмент в такой области, как это отразится на мыслительных и других способностях пациента? Пенфилд был уверен, что какие-то нарушения должны были возникать, но какие? При общении с пациентами после операции он практически не обнаруживал никаких изменений.
Ни один человек в здравом уме не может предположить, что такие большие участки мозга не выполняют никакой полезной функции в организме. Пенфилд хотел, чтобы Хебб узнал, что именно теряет человек вместе с потерей конкретного фрагмента мозговой ткани – даже если эти функциональные потери невозможно обнаружить при обычном общении. В последующие десятилетия исследования показали, что большинство поражений головного мозга действительно влекут за собой потери тех или иных функций. Но что впечатлило Хебба больше всего, так это то, что не терялось: поражения мозга, судя по всему, не вызывали потерю конкретных воспоминаний.
Во избежание недоразумений здесь важно проводить границу между потерей конкретных воспоминаний и потерей конкретных способностей. Большинство из нас знают (читали или непосредственно сталкивались с такими трагедиями, затронувшими членов семьи или друзей), что повреждение мозга может вызвать очень специфические нарушения. Это может быть потеря способности понимать речь (афазия); потеря чувствительности или паралич какой-либо конечности; обвисание лицевых мышц. Иногда функциональная потеря может быть на удивление избирательной: человек лишается способности говорить, но полностью сохраняет способность понимать устную речь. Но это вовсе не то же самое, что потеря отдельных воспоминаний, которую изучал Хебб.
Как высказался по этому поводу сам Хебб, «мы не теряем воспоминание о красных качелях». Если в доме у вашей бабушки была терраса, а на террасе стояли красные качели, мы можем полностью потерять воспоминания о доме и террасе. Но вряд ли случится так, что мы будем помнить яркое изображение террасы и при этом забудем, что на ней были красные качели.
Другими словами, Хебб обнаружил, что пациенты могут терять старые воспоминания (или же терять способность формировать новые воспоминания), но, когда это происходит, все части этих воспоминаний теряются вместе. В обиходе это называется провалами в памяти (если с вами такого никогда не случалось, вы – уникальный человек). Но если уж мы сохраняем в памяти какое-то воспоминание, то помним его практически полностью, со всеми его конкретными составляющими.
Из этого напрашивается вывод, что у конкретных воспоминаний нет четко определенного места в головном мозге. Но как такое может быть? Воспоминания не эфемерные сущности, витающие в некоем эфирном пространстве за пределами нашего физического тела. Они должны быть записаны на каком-то носителе, и этот носитель – головной мозг. Другими словами, все наши воспоминания хранятся в компьютере, который мы называем мозгом, но конкретные воспоминания, кажется, не занимают в нем конкретного физического места.
Концепция нейронных сетей Хебба объясняет не только целостный характер восприятия, но и почему Пенфилд не обнаруживал у своих пациентов существенных нарушений после удаления части головного мозга. Давайте разберемся почему.
Дело в том, что нейронные сети не локализованы в ограниченных зонах, а охватывают обширные области мозга. Такое широкое распределение нервных сетей объясняет, почему конкретные воспоминания не имеют фиксированного местоположения. Если клеточный ансамбль состоит из огромного количества взаимосвязанных нейронов, потеря нескольких из них не причиняет большого вреда. Бо́льшая часть клеточного ансамбля остается взаимосвязанной и функциональной и продолжает репрезентацию восприятия, воспоминания или мысли. Таким образом, конкретные воспоминания не хранятся в конкретном месте: они распределены по всему мозгу. Ответ на вопрос Пенфилда: повреждение нервной сети действительно вызывает функциональные потери, но они настолько малы, что практически неизмеримы (особенно в эпоху Пенфилда). Благодаря нейронным сетям мозг приобретает то, что инженеры называют «отказоустойчивость».
КАК ОБУЧАЕМЫЕ СИНАПСЫ СОЗДАЮТ КЛЕТОЧНЫЕ АНСАМБЛИ
Итак, мы узнали, каким образом клеточные ансамбли создают единое перцептивное целое из отдельных частей и как широко распределенная по мозгу нейронная сеть объясняет устойчивость воспоминаний. Но пока это просто любопытная теория. Что наделяет ее безусловной практической ценностью, так это синапс Хебба – простой и элегантный механизм, объясняющий, как формируются вышеуказанные ансамбли клеток.
Предположим, у нас есть распределенный по мозгу клеточный ансамбль, который обеспечивает и хранит репрезентацию некоего объекта. Клеточный ансамбль – это миниатюрная нейронная сеть. Но почему эта конкретная нейронная сеть имеет отношение к восприятию этого конкретного объекта внешнего мира? Какая связь существует между клеточным ансамблем, который представляет четыре стороны квадрата – то есть квадрат как целостный объект внешнего мира, – и сенсорным опытом восприятия квадратов?
Поскольку наш мир упорядочен, мозг получает входные данные также упорядоченным образом. Видимый мир проецируется на зрительную кору мозга топографически – в виде карты на поверхности мозга. Точнее говоря, в виде карты сетчатки на поверхности зрительной коры. Таким образом, когда на сетчатку падает изображение линии, в зрительной коре активируется группа нейронов, расположенных примерно в виде линии. Зрительная кора – главный портал, через который информация поступает в центры распознавания объектов. Итак, когда сетчатка видит линию, на поверхности коры одновременно активизируются несколько расположенных (примерно) в ряд нейронов. Вы можете догадаться, что происходит дальше: поскольку одновременное возбуждение нейронов усиливает синаптические связи между ними, эти связи в итоге усиливаются настолько, что нейроны «соединяются» в клеточный ансамбль. Другими словами, при первом восприятии линии активизируемые ею нейроны соединены между собой одинаковыми по силе синаптическими связями, но с каждой повторной стимуляцией синапсы внутри этой «линейной» подгруппы нейронов начинают работать все эффективнее. В конце концов, даже активизация одного или нескольких связанных нейронов начинает вызывать возбуждение всего клеточного ансамбля.
После этого кора становится «предрасположенной» к тому, чтобы видеть линии. Даже если сетчатка видит всего лишь часть линии, весь «линейный» ансамбль нейронов возбуждается одновременно и сообщает остальному мозгу, что стимул представляет собой линию. Как правило, такая догадка оказывается верной, потому что линии встречаются в мире гораздо чаще, чем если бы это было случайным событием. По-настоящему случайный, неизбыточный видимый мир – без каких-либо неравномерностей по частоте – был бы похож на снежащий телевизионный экран. Наличие закономерностей в организации мира настраивают мозг на интерпретацию входных сигналов в пользу таких закономерностей.
На следующем уровне сложности четыре линии, встречающиеся в соединенном виде (в виде квадрата), также вызывают одновременное срабатывание нейронов, расположенных примерно в форме квадрата на поверхности зрительной коры. Дальше становится еще сложнее, так как нам нужно перейти от простой двумерной сенсорной репрезентации конкретного квадрата к репрезентации гештальта (обобщенного образа, игнорирующего индивидуальные вариации в деталях) «квадратности», а это уже требует клеточных ансамблей иного порядка. Как это происходит, мы рассмотрим чуть позже, а пока остановимся вот на чем: клеточные ансамбли, представляющие простые геометрические формы, могут объединяться в бо́льшие по размеру нейронные сети, которые выделяют из этих конкретных случаев абстрактное понятие «квадратности». Хебб считал, что такого рода перцептивное обучение (происходящее фактически бессознательно) лежит в основе всего восприятия.
* * *
Усиленное внимание Хебба к восприятию линий не сильно продвинуло нейронауку, потому что в 1960-х гг. были обнаружены свидетельства – хотя и, как оказалось впоследствии, не очень надежные – того, что в мозге есть нейроны с врожденной способностью к распознаванию линий. Казалось, это открытие должно было поставить крест на его теории. Однако базовый элемент этой теории – синапс Хебба, способный обучаться и формировать в мозге клеточные ансамбли, – не умер. Он был забыт почти на десятилетие, после чего впечатляющим образом вернулся вместе с возрождением интереса к нейронным сетям.
ДОНАЛЬД ХЕББ
Я познакомился с Дональдом Олдингом Хеббом – или просто Д.О., как звали его коллеги и студенты, – в 1965 г. Это был худощавый человек среднего роста, сильно хромавший на одну ногу (из-за перенесенного костного туберкулеза одна нога у него была короче другой). Этот дефект слегка изогнул фигуру Хебба, хотя он и старался держаться прямо. У него было квадратное шотландско-ирландское лицо с бледной шотландско-ирландской кожей в обрамлении песочного цвета волос, к 60 годам тронутых сединой[26].
Хебб родился в 1904 г. в Честере в небольшой деревушке, представлявшей собой скопление простых каркасных домов вокруг скалистой гавани на южном побережье Новой Шотландии. Первый Хебб эмигрировал в эту заросшую густыми лесами провинцию из Германии в 1753 г. Впоследствии его многочисленные потомки расселились по всей Новой Шотландии, и недалеко от Честера даже есть городок под названием Хеббвилл. Эти немцы породнились с более многочисленными местными шотландцами, и Дональд Хебб гордился своей шотландской кровью.
Отец Дональда был городским врачом, как и его мать, в девичестве Мэри Олдинг. В 1892 г. она поступила в Университет Дэлхаузи и стала одной из первых женщин в Северной Америке, получившей медицинское образование. По-видимому, именно от нее Хебб унаследовал независимость духа. Его сестра Кэтрин стала известным нейрохимиком и бо́льшую часть жизни проработала в Кембриджском университете.
Семья Хеббов жила в непритязательном доме на главной улице Честера недалеко от гавани. Впоследствии многочисленные Хеббы продолжали приезжать в Честер на каникулы и в отпуск, и сегодня у их семьи по-прежнему есть дом неподалеку от того места. Одной из страстей Дональда было хождение под парусом на небольших яхтах. Как вспоминала жительница Честера, ее брат нанялся к Хеббам обслуживать их небольшой флот и постоянно жаловался на то, что «у этого профессора чересчур высокие требования».
У Хебба действительно были «чересчур высокие требования», и прежде всего он настаивал на оригинальности подходов. Он требовал избегать банальности и искать по-настоящему новые идеи. Недаром на счету его кафедры были такие замечательные – и совершенно неожиданные – открытия, как «центры удовольствия» в головном мозге, области мозга, ответственные за избирательную потерю кратковременной памяти, нейроны пространственной ориентации у крыс и «очаги» эпилептической активности. Все это были настоящие открытия, а не простые усовершенствования или дополнения к уже имеющимся знаниям. Мой сокурсник по Университету Макгилла Джон О’Киф получил Нобелевскую премию за то, что в далеком 1966 г. придумал способ регистрировать активность нейронов в мозге бегающих по лабиринту крыс и обратил внимание на странное поведение некоторых нейронов, которые, как оказалось, помогали крысам ориентироваться в пространстве.
Второе послание, которое Хебб настойчиво пытался донести до всех, касалось важности правильной подачи научного материала. В юности он мечтал стать писателем и потому придавал большое значение стилю. К тому времени, когда я с ним познакомился, Хебб уже не писал беллетристики; он шутил, что уже создал величайший роман своей жизни – «Организацию поведения». Однако он тоннами глотал научно-фантастическую литературу: раз в несколько недель перед его дверью появлялась очередная коробка с книгами, которые он прочитал и теперь предлагал забрать всем желающим.
«Послушайте, – говорил он, – изложение научных идей тоже часть науки. Какая польза другим людям от вашего знания, если оно находится только у вас в голове? Вы должны сделать знание доступным для всех остальных. Неважно, насколько хороши ваши идеи или исследования, – если никто о них не знает, они бесполезны. Вы должны уметь быть убедительными». У меня перед глазами так и стоит картина – Хебб работает над «Организацией поведения» и не перестает себе твердить: «Ты должен продать им эти идеи, чтобы они не канули в Лету». И он гениально справился с задачей: его книга мгновенно стала бестселлером после публикации – не в последнюю очередь благодаря изысканной прозе Хебба, ее ясности и пропитывающей ее научной страсти.
Внешне строгий и дисциплинированный, Хебб был добродушным и заботливым человеком. Секретарем у него в те времена работала необычайно высокая рыжеволосая молодая женщина, прежде – барменша в джаз-клубе. Я не могу рассказать здесь ее историю, но Хебб не побоялся бросить вызов условностям, наняв эту яркую молодую женщину в научное учреждение (разумеется, она знала все лучшие джаз– и блюз-клубы в центре Монреаля).
Хебб применял творческий подход во всем: даже его призывы к творчеству были креативными, то есть довольно необычными. Он твердо верил, что запоминать ненужную информацию вредно, и в качестве доказательства пересказывал шутку о стареющем профессоре ихтиологии, которому каждый раз, чтобы запомнить имя нового студента, нужно было забыть название одной рыбы. Это убеждение зиждилось у него на научном основании: Хебб считал, что информация хранится в памяти благодаря модификации синапсов, поэтому рано или поздно синапсы могут закончиться.
Из этого вытекало побочное следствие, что аспирантов нельзя перегружать формальными курсами. (Первой по важности причиной, почему я выбрал аспирантуру в Университете Макгилла, была возможность учиться у Хебба; второй – отсутствие обязательных курсов.) Единственным жестким условием для получения докторской степени на его кафедре было прослушивание легкого курса по статистике, да и то потому, что этого требовал какой-то высший аттестационный орган. Во всем же остальном вам предоставлялась полная свобода действий.
ПЕРЦЕПТИВНОЕ ОБУЧЕНИЕ
В 1960-е гг. Хебба больше всего интересовало перцептивное обучение. Поскольку под перцептивным обучением могут пониматься разные вещи, давайте разберемся с этим термином подробнее. Первое толкование напрашивается само собой: чем чаще мы выполняем какую-либо сенсорную задачу, тем лучше с ней справляемся. Классический пример – способность локализовать место иголочного укола на коже. Эта чувствительность проверяется легким прикосновением к коже двух иголок, расстояние между которыми постепенно уменьшается. Самое близкое расстояние, на котором человек перестает отличать два укола от одного, называется порогом двухточечной дискриминационной чувствительности. Если вы попросите кого-нибудь проводить вам такой тест каждый день, примерно через неделю вы обнаружите, что стали различать иголочные уколы на гораздо меньшем расстоянии, чем раньше.
Мой любимый пример перцептивного обучения в повседневной жизни – способность слышать музыку. В своей жизни я слушал много музыкальных произведений и могу сказать, что сейчас слышу музыку гораздо лучше, чем в молодости. Я четко различаю текст в полифониях Баха, а также (к сожалению) в песнях стиля хеви-метал. Я не имею в виду, что я стал глубже понимать композиционные тонкости классической музыки или стал знатоком рок-лирики. Речь здесь идет не о понимании, а именно о «тренировке» слуховой сенсорной системы.
Очевидно, что этому есть простое объяснение: благодаря практике человек учится лучше сосредотачивать внимание на своих тактильных ощущения или на музыке Баха. Однако здесь также задействован более глубокий и тонкий механизм перцептивного обучения, который и интересовал Хебба – а именно способность нейронов сенсорной системы настраиваться на восприятие определенного стимула. На самом деле еще в 1949 г., до появления компьютерных наук, Хебб пришел к выводу, что реорганизация нейрональных связей в ответ на закономерности природного мира является не просто сладкой глазурью на торте, а фундаментальной основой всего восприятия.
Мы уже встречались с примером такой закономерности, когда говорили о том, как мы учимся видеть линию. Мир содержит множество визуальных событий, которые характеризуются таким свойством, как избыточность. Посмотрите в окно: в ясный день голубое небо выглядит примерно одинаковым во всех точках. Это и есть избыточность в техническом смысле – одинаковость стимула от точки к точке создает поток одинаковых зрительных сигналов. Это касается и более сложных объектов. Большинство точек на кузове красной машины, скорее всего, будут окружены такими же красными точками, то есть также будут генерировать избыточные визуальные сигналы.
Теперь представьте себе лес, телефонный столб или здание. Все эти три объекта содержат такой важный признак, как прямые линии. Прямая линия также избыточна: если взять любую точку на линии, соседняя с ней точка с довольно большой вероятностью (выше, чем при чистой случайности) будет располагаться на той же линии. В то же время некоторые соседние точки будут находиться в стороне и генерировать другой сигнал. Таким образом, линия по сравнению с ясным голубым небом избыточна лишь частично – в одном направлении, а не во всех направлениях, как в случае неба, – но фундаментального принципа это не меняет. На самом деле в нашем мире очень мало визуальных явлений, которые не являются избыточными. Как уже говорилось выше, по-настоящему неизбыточная визуальная сцена напоминала бы снежащий телеэкран.
Избыточность имеет огромное значение в свете тех требований, которые зрение предъявляет к головному мозгу. Она означает, что мозгу не нужно анализировать каждый пиксель видимой картины. Мозг может выявить в ней имеющиеся регулярные паттерны и закономерности и, опираясь на них, предсказать содержание многих соседних пикселей. Тем самым мозг сэкономит массу времени и сил – ведь закономерности (представленные соответствующими ансамблями клеток) известны ему заранее. Это важнейший общий принцип функционирования биологических сенсорных систем. Мы уже видели его в действии в контексте восприятия краев и столкнемся с ним снова в контексте восприятия таких сложных объектов, как лица.
Размышление об этих закономерностях и навело Хебба на мысль о клеточных ансамблях. Регулярный паттерн (например, линия) одновременно активизирует соответствующую группу нейронов в коре мозга. Повторное восприятие такого паттерна усиливает синаптические связи между этими нейронами, в итоге образующими клеточный ансамбль. Отныне активизация на сетчатке, скажем, трех расположенных в ряд пикселей вызывает возбуждение всего линейного ансамбля – то есть, по сути, мозг предсказывает, что следующие пиксели также будут расположены на этой линии. Конечно, предсказание не всегда будет верным: линия может резко отклониться в сторону, поэтому мозг оценивает вероятность предсказанного события. Но, поскольку видимый мир в основном состоит из регулярных паттернов и закономерностей, предсказания чаще всего сбываются, что с каждым разом повышает эффективность перцептивного прогнозирования.
Говоря иначе, наше восприятие в значительной мере предопределяется нашим предыдущим опытом. Я имею в виду не только то, что вы ожидаете увидеть определенные объекты в определенных контекстах – например, своих братьев и сестер в родительском доме на Рождество, – но и то, что на подсознательном, базовом уровне даже самые простые элементы сенсорных ощущений создаются в той же мере, в какой они реально регистрируются нашими глазами, ушами, кожей или носом.
Один из моих экспериментов в Университете Макгилла как раз демонстрировал примитивную форму таких «созданных» мозгом ощущений. Я изучал феномен, известный как эффект последействия движения. Этот эффект возникает, если в течение длительного времени смотреть на какой-нибудь движущийся узор, а затем посмотреть на тот же узор, но уже неподвижный: у вас создается ощущение, будто узор движется в противоположном направлении. Другое название этого постэффекта – «иллюзия водопада»: если какое-то время смотреть на водопад, а затем перевести взгляд на окружающий неподвижный пейзаж, вам будет казаться, что он движется вверх.
Этот вид оптической иллюзии известен давно. Но мой эксперимент показал кое-что интересное – что эффект последействия движения может сохраняться в течение нескольких дней. Это было важным открытием, поскольку означало, что причина была не в усталости нейронов – как, например, в том случае, когда вы какое-то время смотрите на яркий свет, а затем видите темные круги перед глазами, – а в более стойком изменении восприятия. Я также обнаружил, что этот постэффект был связан исключительно с первоначальным стимулом. Во всем остальном восприятие у испытуемого оставалось абсолютно нормальным, но, когда он смотрел на тот самый узор (прежде двигавшийся, а теперь неподвижный), ему по-прежнему казалось, что орнамент перемещается в противоположном направлении.
Другое мое открытие еще более примечательно: эффект последействия оказывается продолжительным только в том случае, если стимул падает на ту же часть сетчатки, что и раньше. Поскольку визуальные центры мозга организованы в виде карты, вышеописанное изменение зрительного восприятия происходит именно на этом простом уровне сенсорной системы, а не на уровне ожиданий, сознания и т. п. Участники эксперимента не осознавали, что их восприятие как-то изменилось – их зрение было абсолютно нормальным, пока они не обнаруживали странное движение у этого конкретного стимула. У испытуемых менялось – если хотите, «создавалось» – восприятие только этого конкретного объекта, причем очень конкретным и более-менее устойчивым образом в полном соответствии с тем, как это предсказывал Хебб.
Описанный мной феномен был несколько причудливым. Вы «видели» движение тестового стимула, зная, что никакого движения нет (чтобы убедиться в этом, достаточно было сравнить его положение с окружающими стационарными объектами). Это была забавная оптическая иллюзия и не более. Однако позже аналогичный эффект был обнаружен Чарльзом Гилбертом и его коллегами из Университета Рокфеллера, которые изучали дискриминационную чувствительность людей при выполнении такой важной в повседневной жизни перцептивной задачи, как определение местоположения линии в пространстве. Все это говорит о том, что значимое сенсорное восприятие может меняться под влиянием перцептивного опыта.
ДОКАЗАТЕЛЬСТВО КЛЮЧЕВОГО ПОСТУЛАТА ХЕББА
Так сложилось, что идеи Хебба о восприятии и познании не приобрели такой известности, как его постулат о синаптической пластичности (синапс Хебба). Впрочем, это вполне объяснимо: понять и принять эти теории было гораздо труднее, чем концепцию отдельного синапса – реально существующей структуры, которую в деталях можно было разглядеть под микроскопом. Учение же Хебба о восприятии на тот момент не поддавалось проверке. Это стало возможным только в начале XXI в., когда визуализация распределенных нейронных сетей перестала быть несбыточной мечтой.
Сам Хебб не слишком зацикливался на синаптической пластичности. Это был всего лишь принцип, инструмент для построения его теорий восприятия и памяти. Только в XXI в. синапс обрел совершенно конкретные «плоть и кровь». Мы в мельчайших подробностях изучили его строение (сегодня, если хотите, вы можете распечатать модель синапса на 3D-принтере и поставить ее на рабочий стол); мы знаем, какую информацию он передает, с помощью каких веществ, а также знаем десятки типов этих веществ, каждый из которых выполняет конкретную функцию.
В 1949 г. Хебб ничего этого не знал. Синапс лишь недавно стал намного бо́льшим, чем просто постулатом, который должен был быть верным исходя из его объяснительного значения, но не имел прямых доказательств. Проявив чудеса научной прозорливости, Хебб теоретически предположил, что синаптические связи могут модифицироваться и что это свойство может лежать в основе памяти. К началу 1970-х исследователи обнаружили, что изменение силы синаптической связи можно наблюдать в одной из подсистем мозга при помощи довольно простого лабораторного оборудования. Наблюдаемый ими феномен они назвали долговременной потенциацией. (Когда ученые не уверены в интерпретации полученных результатов, они прибегают к таким вот расплывчатым «рабочим» терминам. В данном случае термином «долговременная потенциация» просто описывалось то, что происходило в экспериментах определенного типа. Никто не осмелился назвать это «синаптическим обучением» или чем-то подобным.)
Суть долговременной потенциации (ДП) состояла в том, что многократное возбуждение реального синапса вело к устойчивому усилению синаптической передачи между двумя нейронами в течение нескольких часов и даже дней после стимуляции. Это было именно тем, что постулировал Хебб: нейроны соединяются, если вместе возбуждаются. После этого концепция синапса Хебба и связанные с ней теории породили целую волну исследований. Сегодня поиск в интернете по запросу «долгосрочная потенциация» выдает 13 800 опубликованных статей. В 2000 г. Эрик Кандел получил Нобелевскую премию за открытия, касающиеся синаптической пластичности.
Впервые феномен ДП в лабораторных экспериментах был обнаружен норвежским физиологом Терье Лёмо. В 1966 г. Лёмо опубликовал простое, но точное его описание, за которым в начале 1970-х последовала серия скрупулезно детализированных работ, написанных Лёмо в соавторстве с Тимоти Блиссом и другими. Исследователи изучали проводящие пути в зоне зубчатой извилины в гиппокампе кроликов. Преимущество этой зоны (которая, судя по ее названию, напоминала ранним анатомам форму зуба) в том, что нейроны и их отростки здесь хорошо различимы и доступны для стимуляции, а электростимуляция их аксонов возбуждает известную группу постсинаптических нейронов.
Лёмо и его коллеги обнаружили, что воздействие на пресинаптические волокна (находящиеся перед синапсом) несколькими пачками высокочастотных стимулов вызывало усиление ответа постсинаптических клеток (находящихся после синапса) на протяжении длительного периода времени – нескольких часов или даже дней. Последнее и было самым интригующим, поскольку до сих пор все наблюдаемые нейронные события продолжались в пределах нескольких миллисекунд. Было очевидно, что механизм долговременной потенциации с его устойчивыми изменениями открывал возможность пролить свет на то, как работает наша память.
ВРЕМЕННОЕ ЗАТМЕНИЕ НЕЙРОННОЙ СЕТИ
Я надеюсь, что сумел передать вам, насколько меня восхищает дерзость научного мышления Хебба, который в 1949 г. теоретически описал то, что мы сумели эмпирически подтвердить лишь несколько десятилетий спустя. В 1940-х гг. идея «клеточного ансамбля», или «нервной сети», находилась далеко за рамками воображения ранних нейробиологов, которые были озабочены изучением простых рефлексов – проводящих путей из нескольких нейронов, передающих информацию только в одном направлении. Никто не предполагал существование более сложных сетей (несмотря на то, что на эту возможность намекали рисунки некоторых ранних нейроанатомов). Никому не приходило в голову, что отдельные синапсы могут обладать памятью. И уж тем более никто не пытался объяснить – в одном постулате – нашу способность к распознаванию объектов и устойчивость отдельных воспоминаний при отсутствии части мозга, как это сделал Хебб своей теорией ансамблей клеток.
Кроме того, у Хебба отсутствовали преимущества того концептуального багажа, который имеется в наши дни благодаря развитию компьютерных и информационных технологий. Сегодня большинство мало-мальски образованных людей слышали об искусственных нейронных сетях и машинном обучении. Мы достигли значительного прогресса в этих областях, научились создавать нейронные сети, способные делать такие фантастические вещи, как предсказывать эпидемии до их возникновения или моделировать эволюцию звезд.
У Хебба не было ничего из этого. В 1949 г. исследователи знали, что повреждение определенных частей мозга ведет к нарушению определенных способностей, но никто не осмеливался высказывать настолько детальные предложения о механизме памяти или о связи мозга и восприятия. Хебб совершил прыжок в будущее, сформулировав идеи, в которых мы лишь сейчас начинаем понемногу разбираться.
Как оказалось, он прыгнул в будущее слишком далеко. О его идее нейронных сетей забыли почти на десятилетие. Примерно в начале 1960-х гг. ее подхватила зарождавшаяся область компьютерных наук. Тогда же было сделано несколько интересных изобретений в духе Хебба, наиболее любопытным из которых являлась кибернетическая модель «перцептрон» (подробнее о ней мы поговорим в главе 10). Но к тому времени вышедшая в 1949 г. книга Хебба о клеточных ансамблях уже стала «устаревшей», не будучи написанной на квазиматематическом языке компьютерных наук.
Кроме того, существовало несколько весомых препятствий тому, чтобы извлечь пользу из теорий Хебба. Во-первых, чтобы понять, как работает восприятие, необходимо было ответить на новые, обусловленные знанием о клеточных ансамблях вопросы. Прав ли Хебб в том, что существует континуум от простых до сложных форм восприятия? Как именно соединяются между собой клеточные ансамбли, чтобы обеспечить восприятие более сложных объектов (например, лиц) и далее репрезентацию абстрактных образов? Действительно ли существуют клеточные ансамбли, представляющие не только объекты, но и мысли? И как все это соединено с действием? Но, как я уже сказал, выстраивание таких теоретических конструкций одной только силой разума – колоссальная по сложности задача (намного более сложная, чем экспериментальная работа, где задача разбивается на мелкие части). Кроме того, как и большинство первопроходцев, Хебб старался развить свои идеи по максимуму, насколько хватало полета мысли. Он выдвинул интригующее предположение о реверберирующей (отражательной) активности в клеточных ансамблях, но оно оказалось не таким жизнеспособным, как синапс Хебба. Это и другие расширения его теории только отвлекали внимание от сути.
Вторым серьезным препятствием, которое в конечном итоге заставило многих нейробиологов, включая меня, сосредоточиться на более приземленном биологическом подходе, было отсутствие (даже на горизонте) экспериментальных инструментов для изучения распределенных нейронных систем, чьи элементы (нейроны) по определению разбросаны по всему мозгу. В то время мы были рады и тому, что у нас была возможность наблюдать за одним нейроном зараз. Я описал вам этот трудоемкий процесс, который позволял изучать поведение в лучшем случае полудюжины нейронов в день. Отсюда до наблюдения за целой нейронной сетью, охватывающей обширные области мозга, было все равно что от земли до неба. Поэтому я и многие мои коллеги решили оставить теоретическое поприще когнитивной нейронауки и переквалифицироваться в (нейро-) биологов, занявшись изучением элементарных основ восприятия. Вы уже знаете о результатах нашей работы: благодаря ей мы теперь довольно хорошо представляем, какая информация и в каком виде поступает из глаз в головной мозг. Но как узнать, что происходит дальше? Как ни странно, здесь нам на помощь приходят искусственные нейронные сети.
10 | Машинное обучение, компьютерное зрение и живой мозг
Сегодня почти каждое новое изобретение лишь поначалу принимается с торжествующим криком, а потом он переходит в вопль страха.
БЕРТОЛЬТ БРЕХТ
До сих пор я не высказывался однозначно, но теперь скажу прямо: как и большинство ученых, я считаю, что мозг – это компьютер. Поначалу такого рода сопоставления оставались всего лишь красивой метафорой. В 1980-х компьютеры были настолько несовершенны, что едва могли разобраться с вашей чековой книжкой. Но в последующие годы их вычислительная мощность удваивалась каждые 18 месяцев, как это предсказывал закон Мура, и сегодня от скорости и производительности компьютеров захватывает дух. Также в соответствии с законом Мура хорошие быстрые компьютеры стали относительно дешевы. Конечно, мы, ученые, не можем позволить себе такие же супермашины, которыми пользуются Агентство национальной безопасности (АНБ), Google или ЦРУ. Но нам вполне по карману приобрести достаточно мощный компьютер, способный сравниться если не с мозгом, то хотя бы с маленьким кусочком мозга. Это дало возможность начать создавать модели, которые имитируют мозг или по крайней мере некоторые его функции, и через это лучше понять работу настоящего живого мозга.
Это замечательный прогресс, в котором кроется своя ирония. На заре эпохи компьютерных технологий разработчики создавали зачатки искусственного интеллекта, черпая вдохновение в живом мозге. Но постепенно компьютеры развивались, а люди учились все лучше их обучать, так что в итоге машины в решении некоторых задач превзошли мозг. В результате сегодня не только создатели искусственного интеллекта учатся у живого мозга, но и нейробиологи узнают о мозге много нового благодаря компьютерам и ИИ. Это замечательная улица с двусторонним движением – объединение сил в попытке понять феномен восприятия, и в конце книги мы рассмотрим несколько областей, в которых живой мозг продолжает вдохновлять разработчиков компьютерных систем. Одним из таких направлений, следующим важным этапом в развитии ИИ, является машинное обучение без учителя. Но сначала я расскажу вам о том, что уже умеют делать современные умные компьютеры и что мы можем узнать о нашем собственном мозге благодаря им.
* * *
Лет 20 назад я столкнулся на коктейльной вечеринке с инженером из аэрокосмической компании, которая специализировалась на машинном распознавании образов и, в частности, разрабатывала системы наведения для «умных» бомб. На мой вопрос, используют ли они в своих разработках принципы нейронных сетей, он с ноткой раздражения ответил: «Послушайте, моя задача – создать систему, которая умеет обнаруживать танки среди деревьев. За это мне и платят. Зачем мне вся эта нейронная чушь с ее нечеткой логикой и прочими непонятными вещами? Мне нужно точно знать, как работает моя система, и точка».
Пережитки такого образа мышления сохраняются по сей день. Да, разработчики нейронных сетей не всегда могут сказать, как именно работает их детище, но, несмотря на это, нейронные сети все шире используются для выполнения различных задач, таких как управление автомобилем и т. п. Желание инженеров с их предельно рациональным мышлением детально знать каждый шаг вполне понятно, и в будущем «умные» компьютеры, возможно, помогут им в этом. В конце концов, компьютер всего лишь машина, которая состоит из обычных атомов, и теоретически всегда можно узнать, в каком состоянии находится каждый ее компонент. Поэтому я уверен, что когда-нибудь инженеры в этом разберутся. А тем временем искусственный интеллект будет продолжать совершенствоваться – даже если мы не всегда будем знать, как именно он это делает.
КОМПЬЮТЕРЫ, КОТОРЫЕ СПОСОБНЫ УЧИТЬСЯ
Итак, способность распознавать лица развивается у обезьян благодаря опыту: чтобы в мозге обезьяны появились нейроны, избирательно реагирующие на лица, и участки распознавания лиц, она должна видеть лица. Но как это происходит? Как регулярное восприятие лиц превращается в физическую цепь нейронных связей, идущую из сетчатки в латеральное коленчатое тело, затем в первичную зрительную кору V1 и далее в височную долю, где расположены участки распознавания лиц? На данный момент у нас есть лишь проблески понимания и обрывочные факты с единственной надежной биологической основой – механизмом долговременной потенциации, обеспечивающим усиление синаптических связей по мере практики. Однако мы можем зайти с другой стороны: существует модель, которую мы хорошо понимаем, потому что сами же ее и создали, – и эта модель называется компьютерным зрением.
Дедушкой современных видящих компьютеров (которые вполне можно приравнять к мыслящим компьютерам) был уже упомянутый выше перцептрон. Поскольку перцептрон воплощает в себе базовый принцип машинного обучения, давайте поговорим о нем подробнее.
Первый перцептрон, созданный в конце 1950-х гг., имел простое устройство: он включал набор перцептивных элементов (сенсоров), каждый из которых мог воспринимать один простой параметр, и центр принятия решений, который механистически суммировал входные сигналы и выдавал общий выход. В его основе лежала ранняя версия системы распознавания образов со звучным названием «пандемониум», которая была предложена изобретателем Оливером Селфриджем. Перцептивные элементы в этой системе соответственно назывались «демоны». Каждый демон отвечал за восприятие конкретного признака и мог передавать информацию тремя способами: молчать, шептать «Я вижу немного этого признака» или кричать «Я его вижу!» (одно из значений слова «пандемониум» – адский шум и гам; именно это происходило, когда все демоны принимались одновременно выкрикивать свои сообщения).
Перцептрон способен учиться и становиться «умнее». Предположим, вы хотите научить его распознавать дома. Вы показываете ему изображение дома, в ответ на что все перцептивные демоны сообщают центру решений свои фрагменты информации. Затем вы, выступая в роли учителя, сообщаете центру решений: «Это дом» – на основе чего тот оценивает ввод каждого демона, определяя его ценность для вывода системы в целом (ценность выражается в виде весового коэффициента). Например, демон, отвечающий за обнаружение прямых линий, дал ценный ввод (потому что дома содержат много прямых линий); следовательно, в будущем его голосу будет придаваться чуть больше веса. Хотя сам по себе этот демон не может идентифицировать дом, распознаваемый им признак – прямые линии – имеет непосредственное отношение к изображению дома. И наоборот, демоны, воспринимающие не связанные с домами признаки, дали малоценный ввод, поэтому в будущем их сообщениям будет придаваться меньший вес.
На рисунке ниже показана базовая структура перцептрона. На языке машинного обучения перцептрон называется алгоритмом контролируемого обучения бинарных классификаторов.

Это означает, что перцептрон может принимать решения «да – нет» или в контексте распознавания лиц «Это Бет» или «Это не Бет». Хотя это звучит примитивно, параллельная работа множества таких перцептронов – основа распознавания объектов.
Вот как именно это происходит: изображение в цифровом формате демонстрируется набору демонов, каждый из которых избирательно чувствителен к одному конкретному признаку. Каждый демон определяет присутствие или отсутствие своего признака и отправляет сообщение в центр решений, который собирает входные сигналы от всех демонов. Затем учитель сообщает перцептрону «Это Бет» или «Это не Бет». Центр решений запускает процесс, называемый обратным распространением, суть которого – назначить входному сигналу от каждого демона соответствующий весовой коэффициент: демоны, предоставившие информацию, полезную для идентификации Бет, получают более высокие коэффициенты. На рисунке на предыдущей странице величина весовых коэффициентов показана толщиной линий, идущих от демонов в центр решений (вспомните об усилении синаптических связей между нейронами в мозге). В следующий раз, когда перцептрон увидит изображение лица, центр решений будет суммировать входные сигналы от демонов с учетом их «веса»: одни сигналы будут оказывать большее влияние на конечное решение, другие – меньшее. Эта процедура повторяется множество раз со множеством разных изображений Бет (или любого другого объекта) – и с каждой итерацией перцептрон становится все точнее и точнее. Вот, собственно говоря, и все. Оказывается, даже простейший однослойный перцептрон, такой как показан на рисунке, может достаточно точно научиться распознавать простые объекты, а объединение сотен и тысяч таких перцептронов позволяет создавать системы, способные распознавать лица, водить автомобили и делать другие удивительные вещи.
А теперь рассмотрим конкретный пример. На рисунке на следующей странице показан такой же однослойный перцептрон, но с двумя дополнительными детекторами (демонами в классическом пандемониуме). Задача перцептрона – научиться распознавать букву А.
Каждый детектор настроен на обнаружение одного признака – в данном примере линии определенной ориентации. Когда перцептрону показывается буква А, срабатывают два детектора, чувствительные к линиям, наклоненным справа налево и слева направо, а также детектор горизонтальной линии, реагирующий на перекладину буквы А. Затем центр решений получает от учителя сообщение: «Это буква А» – и через процесс обратного распространения увеличивает весовой коэффициент этих трех детекторов.

Обратите внимание, что на рисунке эти три линии, участвующие в распознавании буквы А, – веса В2, В4 и В5 – более жирные, что призвано подчеркнуть их важность. В следующий раз, когда перцептрону покажут букву, он придаст сообщениям от этих детекторов больший вес.
Теперь предположим, что А-распознающему перцептрону показывают букву Б. В этом случае среагирует детектор вертикальных линий и детектор горизонтальных линий. Но из них двоих для перцептрона имеет значение только сигнал детектора горизонтальных линий. Поскольку два других значимых детектора 2 и 5 молчат, центр решений получает слишком слабый суммарный сигнал и поэтому делает вывод: «Это не буква А».
Следующий важный тест, помогающий понять, как устроено восприятие у машин или мозга, – изменить размер изображения. Мы снова показываем перцептрону букву А, но на этот раз меньшего размера. Как вы помните, машина на основе шаблонов не смогла справиться с этим тестом, потому что маленькая буква А не соответствовала оригинальному шаблону. Но нашему перцептрону достаточно задать всего одно условие: чтобы его детекторы реагировали на линию определенной ориентации независимо от ее размера и местоположения внутри рецептивного поля. В этом случае наш перцептрон получит сигналы, что в этом маленьком символе есть две наклонные линии (слева направо и справа налево) и одна горизонтальная, – и распознает его как букву А.
Как вы обратили внимание, ключевое условие, заданное детекторам в этом примере, полностью соответствует тому, что делают «сложные» клетки зрительной коры. Как и детекторы в нашем гипотетическом перцептроне, эти сложные клетки чувствительны к линиям определенной ориентации независимо от их местоположения в рецептивном поле. Таким образом, эта модель показывает, как со сложной клетки визуальной коры V1 – довольно простого детектора признака, находящегося на относительно ранней стадии визуальной обработки, – начинает выстраиваться распознавание визуальных объектов. Недаром Ян Лекун, один из гигантов в области искусственного интеллекта, сказал, что для него сложные клетки – важный источник вдохновения.
НЕЙРОННЫЕ СЕТИ: БОЛЬШЕ И ЛУЧШЕ
Перцептроны были интересной новинкой, но затем произошло нечто странное: интерес к этому типу искусственного интеллекта почти умер – и возродился лишь четверть века спустя. Впоследствии этот период примерно с 1965 по 1985 г. назвали зимой искусственного интеллекта. В эти годы идею обучающейся машины вроде перцептрона фактически признали бесперспективной. Теперь мы знаем, что это было ошибкой. Сегодня основанный на этих принципах искусственный интеллект догоняет человека. Но почему машинное обучение вызвало такое разочарование?
Во-первых, ИИ в основном являлся плодом эмпирического предположения, не имевшего солидного теоретического фундамента (то есть фундамента, изложенного на языке чистой математики). Это было серьезным недостатком. В те времена в области вычислительной науки работали преимущественно люди с математическим образованием, которые считали недостойным внимания все то, что нельзя было описать математически. На самом деле один ведущий теоретик посвятил целую книгу математическому доказательству того, что простая нейронная сеть такого типа неспособна научиться ничему важному.
Сегодня мы знаем, что он был неправ. Но мы узнали это не с помощью математической теории, а эмпирическим путем – создавая компьютеризированные нейронные сети, которые на деле доказали свою работоспособность.
Вторая причина отсутствия интереса к машинному обучению была чисто практической: в те времена компьютеры были невероятно медленными по сегодняшним меркам и работать с ними было невероятно сложно. Математическая теория оставалась доминирующим инструментом просто потому, что других инструментов как таковых не существовало. Но с появлением все более мощных компьютеров и развитием компьютерных наук растущую роль стал играть элемент эмпиризма, простого метода проб и ошибок. Отныне доказательством могли служить не только математические теории, но и полученные результаты: если нейронные сети работают, значит, они работают, а теория подоспеет, как бы она ни была важна.
Сегодня быстрые и мощные компьютеры и огромные учебные базы данных вывели базовую идею перцептрона на потрясающий новый уровень. На рисунке на следующей странице показана классическая схема современной нейронной сети. Как видите, она состоит из все тех же простых перцептронов, только организованных в несколько взаимосвязанных слоев. Входной слой снабжает входными данными группу из семи перцептронов, а эти семь перцептронов передают обработанные данные на второй слой перцептронов и т. д., вплоть до выхода.

Название «нейронные сети» говорит о том, что эти искусственные модели стремятся имитировать работу живого мозга. Хотя ученые иногда называют их случайными, мы-то знаем, что в действительности они не являются таковыми ни в машинах, ни в живом мозге – просто мы пока не знаем всех правил, по которым они работают. Нам известно, что между искусственными и живыми нейронными сетями есть много общего: те и другие состоят из множества слоев (перцептронов или нейронов). В биологических системах многократная стимуляция одинаковыми входными сигналами приводит к усилению синаптических связей между нейронами. Машинные сети также способны становиться умнее благодаря настройке связей (усилению одних и ослаблению других) между отдельными элементами в процессе обучения.
Как мы увидели, важнейшую роль в машинном обучении играет процесс обратного распространения. Получив ввод от учителя («Это буква А»), современная нейронная сеть производит вычисления в обратном направлении – от выходного слоя к входному, настраивая связи между десятками и даже сотнями скрытых нейронных слоев. (Их называют скрытыми, потому что они, в отличие от входного и выходного слоев, не контактируют напрямую с внешним миром.) Так что теперь перед нейробиологами стоит задача: узнать, делает ли мозг то же самое и, если да, как именно.
* * *
Надеюсь, мои друзья, работающие в сфере машинного обучения, простят меня за столь примитивный рассказ, потому что современные системы искусственного интеллекта гораздо, несравнимо сложнее, чем я вам здесь показал. Взять хотя бы тот факт, что синаптические связи между элементами искусственной сети невозможно усиливать до бесконечности: рано или поздно система достигает предела, выше которого повышение эффективности связей становится невозможным. В этом случае решение состоит в том, чтобы разрешить отрицательную настройку связей – использовать тормозные «анти-Хеббовские» синапсы.
Современные нейронные сети могут принимать любой ввод, выраженный в понятной компьютеру форме: изображения (двумерные матрицы пикселей), объемные объекты (трехмерные матрицы пикселей, называемых вокселями), звуки (последовательности волн давления, оцифрованные надлежащим образом) и даже малограмотную болтовню в социальных сетях. Одним из удивительных ранних примеров стала нейронная сеть, способная обнаруживать начинающиеся эпидемии гриппа. Входными данными для этой сети были написанные разговорным, зачастую неграмотным языком сообщения пользователей на страницах городских форумов в Facebook. В качестве «учителя» использовались отчеты об эпидемиологической обстановке, публикуемые Центрами по контролю и профилактике заболеваний (США). Пройдя обучение, нейронная сеть научилась определять, что в данном городе началась эпидемия гриппа, еще до того, как официальная статистика показывала всплеск заболеваемости. Я не знаю, как именно это делалось, но предполагаю, что среди огромного множества постов на Facebook увеличивалось количество сообщений, связанных со здоровьем, а также частота употребления конкретных слов, таких как «простудился», «провалялся», «разбитый» и т. п. Здесь нет никакой магии; в принципе, такие корреляции мог бы выявить и человек, если бы задался целью проанализировать содержание всех этих миллиардов постов. К счастью, для этого у нас есть компьютеры, а человеческий мозг может заняться более интересными делами.
Как и живой мозг, системы ИИ стремятся быть устойчивыми к повреждению. Если компьютерная нейронная сеть насчитывает мириады соединений, потеря небольшой их части, как правило, незначительно отражается на ее производительности, поскольку каждое соединение содержит лишь крошечную долю от всей совокупности «знаний» сети. Конечно, это неизбежно влечет за собой небольшую функциональную потерю в каком-либо месте, но машина продолжает работать. Как вы помните, такую же удивительную отказоустойчивость нейрохирург Пенфилд обнаружил и у человеческого мозга, который оказался способным функционировать практически нормально даже при удалении значительных участков.
Наконец, как это ни парадоксально, нам вовсе не обязательно знать, что делает каждый скрытый слой в многослойной нейронной сети. Насколько мне известно, никто точно не знает, как нейронная сеть Siri от Apple превращает устную речь в символы на экране айфона. В принципе, это можно узнать: весь секрет кроется в наборе весовых коэффициентов, определяемых для соединений сети. Но, поскольку в этой сети миллиарды соединений, отслеживание того, как именно она распознает фонемы (звуковые единицы), скажем, в произнесенном слове «собака», вряд ли стоит усилий. Если машина работает, значит, она работает.
ТЕРРИ СЕЙНОВСКИ И ГОВОРЯЩАЯ НЕЙРОННАЯ СЕТЬ
Несмотря на «зиму искусственного интеллекта», небольшая группа ученых во главе с Джеффри Хинтоном из Торонто продолжала упрямо разрабатывать концепцию нейронных сетей. Одним из них был Терренс Сейновски, в то время работавший в Университете Джонса Хопкинса. По словам Терри, к счастью, он не читал работу того авторитетного автора, который доказал, что нейронные сети не могут научиться ничему важному. Поэтому он продолжал ими заниматься[27].
Терри Сейновски – выдающаяся и необычная личность. Научную деятельность он начал на знаменитом факультете физики в Принстонском университете, где опубликовал серию теоретико-математических работ о поведении нейронов в головном мозге. После чего неожиданно поступил в аспирантуру на кафедру Стивена Куффлера в Гарварде.
Это был серьезный поворот в научной карьере Терри. Вечный экспериментатор, Куффлер предпочитал изучать нервную систему, в буквальном смысле слова закатав рукава, и не очень любил теоретизировать. Тем не менее из орды соискателей докторской степени он почему-то выбрал Сейновски.
Возможно, Куффлер увидел в нем те же качества, ту же сущностную простоту, которая была присуща ему самому. Сейновски относится к разряду людей, которых сейчас принято называть убергиком (uber-geek, где geek – это фанат, а немецкое слово uber означает «сверх» или «супер»). Он влюблен в свою науку и, кажется, занимается ею днями и ночами напролет. Терри интересует буквально все, он мыслит сразу во всех масштабах, всегда ищет новый угол зрения, скрытый пробел, новаторский путь. Он белая ворона в лучшем смысле этого слова – в науке и в жизни.
Взять хотя бы его манеру одеваться: Терри предпочитает белые рубашки, темные костюмы и классические черные туфли. Большинство ученых жаждут подчеркнуть свою индивидуальность – носят джинсы с биркенштоками, футболки и свитера, водят малолитражки, и многие отращивают бороды. У этого клана свои жесткие нормы и обычаи. Постдок, который приходил на работу на кафедру Куффлера в строгом темно-синем костюме, совершенно явно следовал своим собственным путем. Однажды я пригласил его покататься на яхте, и он явился в элегантных шерстяных брюках и черных туфлях на кожаной подошве.
В общении Терри поразительно раскован: кажется, он говорит все, что думает, и, хотя порой случаются неловкие моменты, это нисколько его не смущает. К тому же у него замечательный гогочущий смех, который слышен издалека. На кафедре нейробиологии не знали, что делать с Терри. В соавторстве с Куффлером он опубликовал интересную и ныне забытую статью о синаптической передаче в простой модели нервной системы. Но эмпирические подходы, культивировавшиеся на кафедре нейробиологии, все же остались для Терри чужими.
После Гарварда Сейновски получил место преподавателя в Университете Джонса Хопкинса и примерно в то же время познакомился с Джеффри Хинтоном, который вместе с Дэвидом Румельхартом и другими изобрел метод обратного распространения ошибки. Этот метод, как мы уже знаем, играет решающую роль в обучении нейронной сети, позволяя уточнять значения синаптических весов в обратном направлении, от выхода через все скрытые слои к входному слою. Сейновски позаимствовал этот замечательный инструмент, чтобы использовать его в своей лаборатории.
Трудясь на поприще клеточной нейробиологии, я ничего не слышал о Терри в начале 1980-х гг. Но в 1985-м я посетил его лабораторию, где он продемонстрировал мне нейронную сеть, которая сама научилась говорить. Я был потрясен.

В нейросеть вводились буквы, по одной за раз. Например, исследователи просили сеть произнести букву c как в слове cat (кот).
Почему эта задача вызывает особый интерес? Потому что английский язык славится своим вариативным произношением, мало подчиняющимся правилам (об этом хорошо знают все иностранцы, которые пытаются его изучать). Например, у нас есть правило, что гласные становятся долгими, если на конце слова есть буква е, как в словах gave и brave. Но в аналогичном случае в слове have буква а произносится совершенно иначе. Мы по-разному произносим букву o в словах mow и cow. Как носители языка, мы этого не замечаем, потому что привыкаем такому с детства. Но для компьютеров все различия подобного рода очень важны – как и для лингвистов, которые не поленились составить длинные перечни правил, исключений из правил и исключений из исключений из правил.
К счастью, они также составили словарь транскрипций, содержащий 20 000 английских слов с их стандартным произношением. Именно этот словарь Терри Сейновски и Чарльз Розенберг использовали в качестве «учителя» для своей нейронной сети. После того как сеть выдавала свой вариант произношения буквы c в слове cat, учитель сверялся со словарем и, если сеть произносила ее как /k/, оценивал ответ как верный. Затем сеть запускала процесс обратного распространения и усиливала связь между фонетическим выводом /k/ и буквой c в слове cat.
Чтобы сделать работу нейросети доступной для внешних наблюдателей, Сейновски и Розенберг (весьма разумно) подключили ее к «диктору» – компьютерной программе, которая преобразует транскрибированные английские фонемы и слова в звуки. Этот последний шаг ничего не давал с точки зрения науки, но позволял убедительно продемонстрировать процесс обучения нервной сети, в котором мог поучаствовать каждый.
До обучения выход нейронной сети, как вы могли догадаться, представлял собой даже не словесную окрошку, а кашу из несвязанных фонем. После нескольких раундов обучения сеть начала выдавать нечто очень похожее на детский лепет: «Га, ба, та». Еще через несколько раундов она начала говорить словами, часто ошибаясь, но постепенно увеличивая количество правильно произнесенных слов. И наконец она заговорила почти на идеальном английском – правильно произнося не только выученные слова, но и любой текст. Самое поразительное, что нейросеть научилась этому, не зная никаких правил английского произношения, только посредством обучения на множестве примеров.
Интересно, что анализ нескольких скрытых слоев показал, что сеть знала даже некоторые словосочетания, хотя в ее архитектуре не были заложены правила английской речи. Создавалось впечатление, будто нейросеть училась говорить по-английски почти так же, как это делают в детстве носители английского языка. Несмотря на то, что мы, англоговорящие, не знаем (за исключением лингвистов) правил английского произношения, мы без особого труда и ошибок читаем вслух по-английски. Другими словами, при изучении английского языка нейронная сеть Сейновски вела себя так же, как человеческий мозг.
Все это делалось на компьютерах начала 1980-х гг. – медленных, как черепаха, по современным меркам. Сегодня компьютеры стали в десятки тысяч раз быстрее, а нейронные сети могут содержать сотни и даже тысячи скрытых слоев. Но, несмотря на возросшую сложность, в основе этих нейросетей лежит все тот же базовый принцип, который использовался Розенбергом, Сейновски, Хинтоном и Хеббом.
Говорящая нейронная сеть поразила не только меня. Сейновски стремительно взлетел на олимп научной славы (и остается там до сих пор). Он стал частым гостем на национальных телеканалах, а метод обратного распространения превратился в стандартный инструмент обучения нейросетей. Вскоре Терри перебрался из Университета Хопкинса в замечательный Институт биологических исследований Солка на побережье Южной Калифорнии, где работает по сей день.
Он по-прежнему носит темные костюмы и ездит на большом черном лимузине немецкого производства. В свои 70 с лишком лет он по-прежнему смеется громким кудахчущим смехом и, несмотря на многочисленные регалии и связанные с этим формальности, остается все тем же убергиком с неискоренимым налетом юношеского задора. Он любит хвастаться своими успехами в работе, но тут нет и тени самолюбования: Терри – скромный человек, который искренне увлечен наукой. Хотя налицо все предпосылки для черной профессиональной зависти, я не знаю никого, кто бы ни обожал Терри Сейновски.
КОМПЬЮТЕРЫ, КОТОРЫЕ УМЕЮТ ВИДЕТЬ
Наверняка вы слышали о видящих компьютерах. Такие компьютеры умеют управлять беспилотными автомобилями и распознавать лица в толпе. Фантасты-паникеры любят рисовать страшное будущее, когда при входе в универмаг Macy’s компьютер будет по изображению с видеокамеры устанавливать вашу личность, изучать ваши покупательские предпочтения, после чего каким-то образом побуждать вас покупать больше товаров.
Могу вас успокоить, что на этот счет не стоит волноваться… пока не стоит. Помните, что капчи[28] все еще служат надежным средством защиты[29]. На самом деле капчи наглядно показывают пределы способностей современных компьютеров (конечно, компьютеры АНБ наверняка могут идентифицировать большинство капчей, но для обычных любительских ботов, пытающихся прорваться на обычные сайты, они не по зубам).
Как бы то ни было, уже сегодня видящие компьютеры удивляют нас своими возможностями – и они продолжают стремительно совершенствоваться. В качестве иллюстрации я расскажу вам о двух способах решения проблемы распознавания лиц, которая, как было сказано в начале этой книги, является Эверестом визуальной нейробиологии.
Лучшие компьютерные системы распознавания лиц сегодня действительно хорошо справляются с этой задачей. Они делают это почти так же хорошо, как и люди, хотя пока не могут сравниться с человеческим мозгом в компактности и энергоэффективности. Давайте рассмотрим два противоположных подхода. Первый – основанный на правилах, это означает, что он построен на выполнении серии строго заданных аналитических шагов. Такой подход обычно первым приходит в голову большинству людей (включая того самоуверенного аэрокосмического инженера, о котором я упомянул в начале главы). Для краткости мы будем называть основанный на правилах метод «глупым», хотя его варианты могут отличаться значительной сложностью.
Второй подход основан на машинном обучении и все больше начинает напоминать работу человеческого мозга. На данный момент кажется, что за этим подходом будущее. Именно он приводит в ужас защитников конфиденциальности. Назовем все версии на основе ИИ «умными» методами – потому что они имитируют работу нашего мозга, а я считаю мозг умной системой, – и дальше я сосредоточусь главным образом на них. На сегодняшний день именно умные методы доминируют в области распознавания лиц.
Задача алгоритма распознавания лиц состоит из двух основных шагов: сначала определить наличие лица, а затем установить, кому оно принадлежит. Первая задача называется обнаружением лица, вторая – идентификацией. Умные и глупые методы начинают с одинаковой исходной проблемы: им нужно проанализировать визуальную сцену со множеством различных объектов – например, в отделе мужской одежды в универмаге Macy’s, чтобы определить, есть ли там лица, и изолировать эти лица для дальнейшего анализа.
Но прежде, чем искать лица, компьютеру нужно подготовить изображение, сделать его максимально четким (для себя). Эти шаги, предшествующие запуску алгоритма обнаружения лиц, собирательно называются предварительной обработкой[30]. Существует множество разных способов улучшить четкость изображения, как это знает каждый, кому приходилось работать с Adobe Photoshop. Вот два примера. Во-первых, большинство естественных сцен имеют неравномерную освещенность: на открытом воздухе солнце создает тени и блики; в отделе мужской одежды Macy’s то же самое делает подсветка витрин. Благодаря особым свойствам нашего зрительного анализатора мы не замечаем различий в освещенности, чего нельзя сказать о цифровой камере в смартфоне или компьютере. Их педантичный цифровой анализатор воспринимает один и тот же по-разному освещенный объект как два разных объекта. Следовательно, первая операция по нормализации – «выравнивание» яркости. Компьютер вычисляет среднюю яркость всего изображения (иногда используя для этого мудреные средне-подобные величины) и затем корректирует яркость различных участков изображения так, как если бы вся сцена была равномерно освещена одинаковым источником света. Во-вторых, в большинстве случаев выполняется выделение краев, потому что, как мы уже не раз говорили, именно края несут основную информацию об объектах.
Подготовив изображение, компьютер может приступить к поиску лиц. Существует несколько способов это сделать. Один из наиболее любопытных, который отчасти копирует работу нейронных сетей в зрительной коре мозга, называется методом HOG (Histogram of Oriented Gradients), или гистограммой направленных градиентов.
В нашем случае градиент – это переход между ярким и менее интенсивным цветом, то есть мы имеем участок изображения (край), одна сторона которого яркая, а другая – тусклая. Другими словами, градиент показывает не только наличие края, но и его направление – внутрь или наружу. Компьютер измеряет все возможные градиенты на изображении, определяет их направление и отображает их все в виде карты.
На рисунке на следующей странице представлен пример такого редуцированного HOG-изображения. Чтобы его получить, алгоритм разбил исходное изображение на небольшие квадраты, 16×16 пикселей каждый. В каждом квадрате он вычислил градиенты по всем основным направлениям (вверх, вниз, по диагонали и т. д.) и затем заменил изображение в каждом квадрате его сжатой репрезентацией – обычной линией, показывающей направление края, наиболее выраженное в данной ячейке.
На самом деле здесь вы видите своего рода усредненную, универсальную HOG-структуру лица, созданную на основе множества изображений лиц. Такой шаблон удобно использовать для обнаружения лиц в толпе. Для этого компьютер преобразует каждый участок изображения толпы – каждый квадрат пикселей, размер которого примерно соответствует размеру лица, – в HOG-карту. После чего перемещает универсальный HOG-шаблон по всему преобразованному изображению, сравнивая локальные HOG-квадраты с шаблоном. Большинство квадратов будет содержать беспорядочную мозаику краев. Но некоторые из них приблизительно совпадут с HOG-шаблоном и, следовательно, будут идентифицированы алгоритмом как лица. Конечно, это не позволяет нам идентифицировать, кому именно принадлежит лицо. Но зато мы получаем изображение лица, которое можно преобразовать в стандартный формат и использовать как ввод для последующих шагов классификатора, которому гораздо проще работать с такими входными данными, чем с необработанными изображениями. Таким образом, алгоритм взял беспорядочную мешанину пикселей – отображающую визуальную сцену с толпой – и вычленил из нее несколько локальных массивов пикселей, которые предположительно являются лицами.

Итак, мы имеем нормализованное изображение с локализованными на нем лицами. Посредством нескольких дополнительных операций изображения лиц переводятся в стандартный формат (например, обрезаются по контуру, чтобы исключить окружающий фон, – такие тонкие рамки вокруг лиц вы можете видеть в научно-фантастических фильмах). Теперь компьютер готов перейти к следующему шагу – попытаться определить, кому принадлежит лицо.
Если выйти на Таймс-сквер и спросить у случайных прохожих, как компьютер распознает лица, большинство из них ответит примерно следующее: «В компьютер закладывают правила об основных параметрах лица. Например, он измеряет расстояние между глазами или высоту лба. А затем сравнивает эти параметры неизвестного лица с параметрами лиц, которые есть в его базе данных».
Действительно, системы распознавания лиц на основе фиксированных правил существуют. Например, разработанный Мэтью Терком и Алексом Пентлендом алгоритм использует расчет так называемых собственных векторов для главных компонентов лица. Но на сегодняшний день большинство систем распознавания лиц используют машинное обучение. Разумеется, ситуация может измениться, и в будущем правиловые алгоритмы вполне могут вернуться на сцену (вспомните о моем предсказании через десяток лет). Но пока этого не произошло, мы сосредоточимся на обучающихся нейронных сетях.
Давайте посмотрим, как работает простой алгоритм распознавания лиц на основе машинного обучения, который входит в пакет инструментов MATLAB. Это полезное упражнение, потому что, как выяснилось, человеческое восприятие функционирует во многом похожим образом. Первые шаги по преобразованию изображения лица в стандартный формат не отличаются от тех, что описаны выше для основанного на правилах алгоритма. А дальше происходит вот что:
● Огромное множество обработанных изображений лиц (равномерно освещенных, с выделенными краями, анфас) вводится в многослойную нейронную сеть. Это этап машинного обучения. Каждое лицо имеет свое имя: «Дик», «Джейн», «Билл» и т. д. И для каждого изображения лица вы сообщаете машине: «Это Билл» или «Это не Билл».
● Нейронная сеть, как и простой маленький перцептрон, использует обратное распространение для настройки весов отдельных соединений. Те соединения, которые подают самый сильный сигнал, когда учитель сообщает, что «это Билл», усиливаются. Разница лишь в том, что мощная система использует целый массив простых перцептронов, организованных в скрытые слои. В процессе обратного распространения происходит настройка всех этих скрытых слоев вплоть до входного.
● После обучения нейронную сеть нужно протестировать: в сеть вводится другое изображение Билла, которое не входило в обучающий набор. Усиленные синаптические связи должны среагировать на специфические черты лица Билла, благодаря чему центр решений должен получить сильный суммарный сигнал и выдать ответ: «Это Билл».
Очень большая нейронная сеть, обученная на большом количестве лиц, становится очень умной: она способна узнать лицо Билла с разных сторон, ярко освещенным или затененным, с воротничком белой рубашки или воротом красной футболки. В реальной жизни для обучения нейронных сетей распознаванию лиц используются гигантские базы данных. Раньше такими базами служили, например, архивы фотографий с водительских прав, содержащие миллионы идентифицированных изображений.
Самое удивительное, что мы не знаем, как именно нейронная сеть распознает лицо Билла. По оттенку кожи? Соотношению высоты и ширины лица? Форме носа? Ямочках на щеках? Старым послеугревым рубцам? По всему вышеперечисленному? Все это скрыто в глубинных слоях нейронной сети с их сотнями тысяч соединений.
В главе 11 мы подробнее поговорим о том, чем биологическая зрительная система похожа на видящие компьютеры. Я утверждаю, что обучаемые синапсы играют ключевую роль в основанных на нейронных сетях зрительных системах на всех этапах процесса восприятия – от сетчатки (входного слоя) до высших уровней, где происходит распознавание объектов. Но сразу предупреждаю и подробнее объясню это в главе 13 (внимание, спойлер!): распознавание объектов в биологической зрительной системе происходит вовсе не так, как это делает вышеописанный алгоритм MATLAB. С точки зрения живого мозга это слишком глупый способ. Перцептронам нужен учитель, который будет контролировать их обучение, сообщая им: «Это Билл» и «Это не Билл». Живой мозг способен учиться без внешнего учителя (отдельная важная тема, о которой мы также поговорим подробнее). Но в том и другом случае в основе, по сути, лежит одинаковый ключевой принцип – сети нейронов, связанные модифицируемыми синапсами Хебба.
11 | Как работает зрение
Большинство фундаментальных научных идей по существу просты и могут быть выражены языком, понятным каждому.
АЛЬБЕРТ ЭЙНШТЕЙН
Итак, пришло время наконец-то дать ответ на вопрос, с которого началась эта книга: как родители узнают своего ребенка на детской площадке? При всей кажущейся простоте этот вопрос – как происходит распознавание объектов в мозге – является одной из сложнейших проблем нейробиологии. Картина, нарисованная мной, отличается от той, что представлена в большинстве учебников, которые явно или неявно постулируют существование фиксированной иерархии инкрементальных шагов, каждый из которых ведет к следующему шагу более высокого уровня. На самом деле, как показывают последние исследования, наш зрительный анализатор практически от начала и до конца основан на гибких механизмах нейронной пластичности, обучающихся по правилам нейронной сети.
Для начала я хотел бы показать коммутационную схему зрительного анализатора. К счастью, мне не пришлось составлять ее самому: эта трудоемкая работа уже проделана Даниэлем Феллеманом и Дэвидом Ван Эссеном из Университета Вашингтона в Сент-Луисе, которые составили схему связей в зрительной системе приматов. Как подчеркивают ее авторы, здесь отражены только самые основные соединения. Прямоугольники – это области мозга. Линии – аксональные проводящие пути между ними. Мы, нейробиологи, любим показывать эту схему как наглядное доказательство ужасающей сложности мозга. Помимо прочего, тем самым мы отчасти пытаемся оправдаться за то, что мы до сих пор не разобрались в его устройстве. Еще раз повторю: на этой схеме показаны соединения только между наиболее крупными областями мозга. Если бы мы попытались показать все соединения между нейронами, этих линий было бы в миллионы раз больше. В таком масштабе вместо коммутационной схемы вы бы увидели квадрат густо-черного цвета.

Давайте начнем с широкого, базового представления о том, как устроена наша зрительная система. Конечно, сегодня еще слишком многое остается неизвестным, а высшие зрительные центры изучены лишь в самых общих чертах. Но в ожидании того, когда экспериментаторы наконец-то доберутся до отдельных соединений на уровне нейронов, мы можем выделить ключевые фундаментальные принципы организации нашего зрения. Вот они:
1. Зрительные системы не являются нейтральными, беспристрастными регистраторами входных сигналов. Они искажают (модифицируют) свои ответы на каждом уровне, приводя их в соответствие с закономерностями видимого мира.
2. Иногда это свойство встроено в генетический код, но во многих случаях оно является результатом обучения нейронной сети. Это касается всего – от базовых закономерностей, таких как края и линии, до восприятия сложных объектов, таких как лица.
3. Грубые соединения между зрительными областями мозга образуются при помощи сигнальных молекул – таких же, какие используются природой, например, чтобы обеспечить развитие печени или рук в ходе внутриутробного периода. Эти молекулы помогают аксонам зрительных клеток найти путь к областям-мишеням в головном мозге и сформировать примерную топографическую карту поля зрения в каждой из них. Более тонкие нейрональные связи, лежащие в основе восприятия конкретных объектов – распознавания объектов, – создаются благодаря механизму нейронной пластичности.
ЗРИТЕЛЬНАЯ СИСТЕМА МОЗГА КАК НЕЙРОННАЯ СЕТЬ
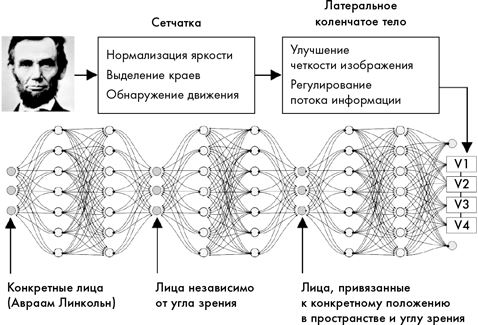
В предыдущих главах я изложил вам основные факты о зрительном анализаторе, которые мы узнали экспериментальным путем к сегодняшнему дню. Вкратце повторим их:
1. Сетчатка предварительно обрабатывает изображение, разбивая его на множество независимых репрезентаций.
2. Сетчатка передает зрительную информацию в латеральное коленчатое тело, которое повышает четкость рецептивных полей и регулирует поток информации, идущий в кору.
3. Первичная зрительная кора (V1) трансформирует рецептивные поля; большинство нейронов V1 лучше всего реагируют на края определенной ориентации.
4. В зрительных зонах V1 и V2 многие клетки освобождаются от жесткой настройки на конкретное положение стимула: эти так называемые сложные клетки реагируют на края определенной ориентации, которые могут находиться в более широкой области видимого пространства. Это шаг к абстрагированию от точного визуального ввода.
5. Следующие зрительные зоны, V3 и V4, содержат нейроны с разнообразной избирательной чувствительностью – например, к цвету, движению или глубине. Они передают информацию в зрительные области в височной коре.
6. Нижняя височная кора состоит из мозаики отдельных зон, чувствительных к самым разным вещам. Некоторые из этих зон специализируются на распознавании лиц.
7. В направлении от задней к передней части височной доли участки распознавания лиц снижают свои «позиционные» требования: их нейроны постепенно приобретают пространственную инвариантность, то есть реагируют на лица независимо от их точного местоположения или ориентации в пространстве.
8. В средней височной коре и еще более высоких областях коры находятся клетки, которые реагируют только на изображение конкретного человека или объекта независимо от его местоположения в поле зрения и от угла зрения.
Раньше ученые считали, что большинство этих шагов жестко запрограммированы в мозге в виде структуры с фиксированными связями. Но, как мы теперь знаем из многочисленных исследований, эта система оказалась гораздо более пластичной. В следующих разделах мы еще раз пройдемся по зрительной системе от сетчатки до высших корковых областей, но на этот раз сосредоточим внимание на ее сетеподобном характере, а также на значительной пластичности и обучаемости. Нас также будут интересовать аспекты, которые делают естественное человеческое зрение похожим на ведущую форму компьютерного зрения.
СЕТЧАТКА
Компьютерное зрение обычно начинается с этапа предварительной обработки или нормализации изображения, в ходе которого беспорядочное естественное изображение преобразуется в более простое и пригодное для дальнейшей обработки. То же самое, по сути, делает и сетчатка: она улавливает свет и преобразует вывод своих светочувствительных клеток – палочек и колбочек – в набор сигналов, с которыми может работать остальная часть зрительной системы. Первым делом сетчатка нормализует вывод с фоторецепторов, выравнивая огромные вариации интенсивности света, которые характерны для земной реальности. Это гораздо важнее, чем мы думаем. Значения сигналов палочек и колбочек разнились бы в сотню миллиардов раз темной безлунной ночью и ярким солнечным днем, если бы их вывод не нормализовался сетчаткой. Ни отдельные нейроны, ни весь мозг, ни даже компьютеры не способны справиться с таким гигантским диапазоном входных сигналов.
Сетчатка сжимает этот диапазон, так что при любой освещенности ее выходные сигналы об интенсивности света варьируются всего примерно в десять раз. Что еще удивительнее, она центрирует этот узкий диапазон относительно средней яркости окружающего освещения на данный момент[31]. Мы осознаем этот процесс настройки, только когда внезапно выходим из темноты на свет или наоборот: мы временно ослеплены ярким светом или ничего не видим в темноте, пока сетчатка не адаптируется к новому уровню яркости. Второй вид предварительной обработки изображения, который выполняет сетчатка, – это начальное выделение краев (при помощи латерального торможения) и обнаружение движения, о чем мы говорили в главе 4.
В чем смысл этого начального этапа обработки изображений? В компьютерах практически любой алгоритм машинного зрения начинается с набора операций, цель которых – уменьшить вычислительную нагрузку на последующие этапы обработки, будь то анализ на основе правил или с использованием нейронных сетей. Например, природа на протяжении миллионов лет усвоила ключевой урок: важно то, что движется, – и воплотила это знание в сетчатке в виде чувствительных к движению ганглионарных клеток.
ЛАТЕРАЛЬНОЕ КОЛЕНЧАТОЕ ТЕЛО (ЛКТ)
В позднем пренатальном периоде аксоны ганглионарных клеток сетчатки уже достигают ЛКТ. Там они соединяются с нейронами, но делают это неточно: окончания ганглионарных клеток разветвляются на множество веточек, которые охватывают довольно обширные области и соединяются со многими нейронами ЛКТ. Если бы такая ситуация сохранялась и в зрелом возрасте, наше зрение было бы размазанным из-за перекрывающих друг друга проводящих путей. Но благодаря механизму синаптического усиления аксоны сетчатки все лучше достигают своей цели.
Вкратце это работает следующим образом: генетически запрограммированные молекулярные сигналы направляют аксоны сетчатки к ЛКТ, где те формируют грубую топографическую карту. Аксоны, которые одновременно возбуждают свои постсинаптические нейроны, – то есть аксоны, идущие из одного глаза, – усиливают синаптические связи со своими нейронами-мишенями в ЛКТ. Постепенно разветвленные аксоны уточняют свои мишени, так что одна группа нейронов ЛКТ становится чувствительной к входящим сигналам от правого глаза, другая – от левого. Страйкер и Шатц убедительно доказали это посредством обстоятельно проведенных и воспроизводимых экспериментов, что стало очередным важным шагом в исследовании зрительной системы.
ПЕРВИЧНАЯ ЗРИТЕЛЬНАЯ КОРА (V1)
Начиная с этого места мы можем рассматривать этапы зрительного анализатора как слои нейронной сети. Давайте возьмем процесс распознавания объектов и посмотрим, как каждый из его этапов мог быть создан мозгом с использованием того, что сегодня мы называем правилами машинного обучения.
Нейроны ЛКТ проецируют аксоны в первичную зрительную кору, где появляются нейроны, чувствительные к ориентированным краям. Но как нейроны V1 создают простые ориентированные рецептивные поля из неориентированных полей нейронов ЛКТ?
Дело в том, что в нейроне V1 сходятся аксоны нескольких нейронов ЛКТ, причем сходятся весьма специфическим образом:[32] рецептивные поля этих нейронов ЛКТ располагаются на сетчатке в ряд и их аксоны выстраиваются в нейроне V1 таким образом, чтобы отражать это линейное расположение. Если схематично зарисовать ответы нейрона V1 на точечное пятно света, карта его рецептивного поля будет выглядеть так, как показано на рисунке на следующей странице слева. Но на отдельные точечные стимулы он реагирует очень слабо; сильнее всего он возбуждается при виде края, ориентированного вдоль ряда плюсов (областей возбуждения).

Рецептивные поля нейронов ЛКТ – это маленькие кружки на рисунке слева. Аксоны клеток ЛКТ сходятся в одном нейроне первичной зрительной коры. Одни из них идут от on-клеток и являются возбуждающими (обозначены плюсом); другие – от off-клеток и являются тормозящими (обозначены минусом). Если сетчатка стимулируется краем, точно ориентированным под соответствующим углом (рисунок справа), все возбуждающие входы активируются, а все тормозящие молчат.
Корковый нейрон имеет вытянутое рецептивное поле с областью возбуждения, граничащей с областью торможения. Оптимальный стимул для этой клетки показан на рисунке справа: светлая область, граничащая с темной. Именно это мы и называем ориентированным краем. На светлой стороне края входы четырех нейронов ЛКТ суммируются, когда край имеет соответствующую ориентацию.
Как уже не раз отмечалось, линии и края очень важны, поскольку они являются доминирующими носителями информации о естественной визуальной среде. Это объясняется тем, что наш мир состоит из объектов, а края объектов определяют их границы, отделяя от всего остального. Иногда края бывают прямыми, как стволы деревьев; иногда изогнутыми, как у камней, но кривая – это просто набор крошечных прямых линий. Таким образом, информация о краях составляет значительную часть общего ввода в зрительную систему мозга, что неизбежно влияет на организацию ее синаптических связей.
Нам уже известен механизм, посредством которого нейронная сеть, обучаемая на примерах из естественного мира, может трансформировать ввод от круглых неориентированных рецептивных полей в чувствительные к линиям элементы – то есть создавать «простые» корковые клетки. Представим группу клеток ЛКТ, сходящихся в нейроне V1. Когда четыре расположенные в ряд клетки ЛКТ стимулируются краем соответствующей ориентации, они одновременно возбуждаются и вызывают возбуждение своего коркового нейрона. Это приводит к усилению синаптических связей между этой линией клеток ЛКТ и нейроном-мишенью – и, как следствие, к относительному ослаблению связей этого нейрона с другими клетками ЛКТ.
Описанный механизм был протестирован много лет назад на искусственной нейронной сети, которую обучали путем простой демонстрации большого количества изображений естественного мира (это был случай так называемого обучения без учителя). Последующий анализ выходного слоя сети показал, что она научилась распознавать прямые линии – то есть в процессе обучения компьютер создал свою версию простых клеток. Это неудивительно: поскольку видимый мир непохож на случайно снежащий телеэкран, практически любая зрительная система – естественная или искусственная – уже на ранней стадии будет включать функцию обнаружения краев.
ЗРИТЕЛЬНАЯ ЗОНА КОРЫ V2: СЛОЖНЫЕ КЛЕТКИ
Сложные клетки, как и их простые собратья, избирательны в отношении ориентации, но имеют более обширное рецептивное поле и, как следствие, менее привязаны к точному положению ориентированного края в пространстве. Можно сказать, что эти клетки выполняют базовую форму генерализации: они выделяют общий признак – край определенной ориентации, абстрагируясь от конкретного набора зрительных пикселей, стимулируемых на сетчатке.
Предположительно сложная клетка создается так же, как и простая: посредством конвергенции нейронов предыдущего уровня, в данном случае простых клеток, в нейроне более высокого уровня. Каждая простая клетка чувствительна к ориентированному краю, находящемуся в строго определенном месте поля обзора. Схождение множества простых клеток с ограниченными локальными полями в одном нейроне создает сложную клетку, которая реагирует на край данной ориентации на всем пространстве их общего рецептивного поля.

Сложные клетки сосредоточены в зрительной зоне V2, но также присутствуют и в первичной зрительной коре V1. Поэтому сделанное мной здесь разделение призвано всего лишь подчеркнуть тот факт, что сложные клетки, судя по всему, создаются путем иерархической конвергенции простых клеток, как это предположили Хьюбел и Визель. Этот используемый мозгом способ создания сложных клеток из простых лег в основу одной из ведущих современных форм машинного обучения.
ЗРИТЕЛЬНЫЕ ЗОНЫ КОРЫ V3 И V4
Многочисленность и разнообразие связей между зонами V1, V2, V3 и V4 не позволяет рассматривать их как жестко обособленные анатомические структуры, а также затрудняет понимание того, каким образом происходит формирование последующих рецептивных полей из предыдущих. Скорее речь идет о сетеобразной структуре.
Изучение активности нейронов в зонах V3 и V4 показывает, что те реагируют на различные признаки. Один из достаточно хорошо изученных – кривизна. Первоначально эти клетки были описаны Хьюбелом и Визелем как клетки, реагирующие на концы линий (end-stopped cells). Эти клетки чем-то похожи на сложные клетки – предполагается, что они создаются посредством иерархической конвергенции сложных клеток, – но обладают дополнительным свойством: они предпочитают не просто края, а края определенной, фиксированной длины. Однако последующие исследования показали, что это свойство также можно рассматривать как чувствительность к кривизне. Помимо них в зонах V3 и V4 содержатся клетки с другими специфическими видами чувствительности: так, некоторые клетки в V4 чувствительны к цветам, а в V2 есть клетки, чувствительные к углам. Честно говоря, ученые пока имеют весьма смутное представление о том, что происходит внутри этих зрительных зон.
По аналогии с искусственными нейронными сетями зоны V1, V2, V3 и V4 могут функционировать как промежуточные «скрытые слои» зрительной сети. Именно это может объяснять, почему поведение их нейронов – несмотря на все усилия исследователей – так трудно классифицировать. Как мы увидели в главе 10, скрытые слои последовательно соединяют несколько нейронных сетей, таким образом значительно увеличивая их вычислительную мощь. Они называются «скрытыми», потому что не взаимодействуют напрямую с внешним миром, а только с соседними слоями. Понять, что делают эти скрытые слои, нелегко даже в собственноручно сотворенных нами системах ИИ. Ситуация осложняется тем, что нейроны в одном скрытом слое необязательно должны делать одно и то же. Да, у нас есть основания полагать, что нейроны в зонах V3 и V4 выполняют более сложные функции по выделению признаков, чем клетки в зонах V1 и V2. Можно сказать, что по своей сложности функции этих скрытых слоев находятся в промежутке между функциями зон V1 и V2 и функциями областей распознавания объектов в височной коре.
ВИСОЧНАЯ КОРА
Если не вдаваться в детали, обработка зрительной информации в височной коре построена по иерархическому принципу в направлении от задней к передней части коры: более простые признаки выделяются в задней части, ближе к первичной зрительной коре V1, а распознавание сложных объектов происходит ближе к передней части (лобной доле мозга). Разумеется, это сильно упрощенное представление (вспомните вышеприведенную анатомическую «коммутационную схему» с ее паутиной прямых и обратных связей), но достаточно близкое к реальности, чтобы быть полезным для понимания системы обработки изображений в мозге.
В височной доле существует по крайней мере шесть участков распознавания лиц, соединенных между собой аксональными связями. У этих участков есть свои формальные названия, указывающие на соответствующие анатомические подразделения височной доли, где они расположены. Но эти названия слишком сложны для неспециалистов и вызывают разногласия у экспертов, поэтому я взял на себя смелость свести их в более обобщенную номенклатуру. Не вдаваясь в детали, я расскажу о задней, центральной и передней частях нижней височной коры.
Продолжая наш разговор, мы можем рассматривать эти шесть участков распознавания лиц, распределенных по коре височных долей, как скрытые слои нейронной сети, предназначенной для идентификации конкретных визуальных объектов. Самое примечательное, что эти зоны коры являются в определенной мере универсальными распознавателями – они не запрограммированы генетически на распознавание лиц, а приобретают эту специализацию в процессе обучения (например, эксперименты Ливингстон и ее коллег показали, что у обезьян, которые с детства не видели лиц, эти участки отвечают за распознавание рук).
Можно сказать, что скрытые слои нейронной сети в направлении от задней к центральной части височной коры выделяют все более сложные наборы признаков лица. Скрытые слои в задней части получают входные данные от зон V1–V4 и используют их для обнаружения таких компонентов, как овал лица, нос, подбородок, волосы и особенно глаза. Нетрудно представить, как сложная избирательная чувствительность предыдущих слоев – избирательность к кривым, углам и т. д. – помогает обнаруживать вышеуказанные компоненты лица. В магии распознавания лиц гораздо меньше магического, чем может показаться, потому что лицо – не случайный набор пикселей. Но детальная механика этого процесса нам (пока) неизвестна: как это часто бывает с нейронными сетями, чем именно занимается каждый скрытый слой, остается загадкой.
Участки распознавания лиц в задней и центральной частях височной коры, по всей видимости, компонуют выделенные признаки лица в простую репрезентацию – своего рода «протолицо». Эксперименты Цао и ее коллег показали, что эти признаки предположительно являются относительно простыми, например, такими как соотношение высоты и ширины лица, расстояние между глазами и т. д. Еще раз повторю, что пиксели в изображении лица расположены не случайным образом. Два темных пятна, соответствующих ноздрям, обычно идут парой, а ниже них находится линия пикселей, соответствующая рту. Отдельные элементы лица связаны между собой в реальном мире – и становятся связанными в мозге в виде клеточного ансамбля.
Нейроны в указанных участках чувствительны к изображениям реальных биологических лиц, но их легко обмануть символическими лицами – овалами с двумя точками вместо глаз и короткими прямыми линиями вместо носа и рта. Исследования Цао и ее коллег позволяют сделать вывод, что некоторые находящиеся здесь нейроны могут математически суммировать отдельные компоненты и делать вывод о степени подобия лицу. Играя с различными комбинациями компонентов, они обнаружили, что, например, простое лицо без одного глаза вызывает у таких нейронов менее сильный количественно измеримый ответ, чем такое же лицо с двумя глазами. Но даже эти продвинутые нейроны требуют, чтобы изображение лица попадало в строго определенное место их рецептивного поля – аналогично простым клеткам в зоне V1, которые реагируют только на строго локализованные ориентированные края.
Логично предположить, что задняя и центральная части височной коры поставляют зрительную информацию в следующий нейронный слой – переднюю часть, которая является пространственно-инвариантным распознавателем лиц. Многие нейроны в передних участках допускают относительную свободу в отношении расположения лица в пространстве, а также представляющего его набора пикселей. Механика этого процесса также детально неясна, но можно предположить, что она напоминает происходящее в аналогичных искусственных нейронных сетях. Установлено, что некоторые находящиеся здесь нейроны способны распознавать не только прямое, но и зеркальное изображение лиц в обширном рецептивном поле. Зачем нужно распознавание зеркальных изображений, пока неясно. Правдоподобное предположение состоит в том, что эти клетки могут быть промежуточным скрытым слоем – звеном на пути к достижению полной пространственной инвариантности. И действительно, в самом переднем участке есть такие продвинутые нейроны, которые реагируют на лица независимо от их расположения в пространстве.
Наконец, что еще более примечательно, в близлежащей области коры обнаружены нейроны, реагирующие на конкретные лица. Это означает, что в нашем мозге могут иметься клетки и микросхемы – части клеточных ансамблей, помогающие нам узнавать всех знакомых нам людей: членов семьи, друзей, коллег и т. д. Предположительно выходные данные из пространственно-инвариантной нейронной сети служат входными данными для нейронной сети еще более высокого уровня, которая учится распознавать конкретных людей. Но как эти клетки интегрируются в целостную систему, мы пока можем только гадать[33].
Таким образом, этапы обработки зрительной информации в височной коре можно представить как серию из пяти видов событий. Во-первых, нейронная сеть учится распознавать компоненты лица. Во-вторых, из этих выделенных компонентов лица – глаз, носа, рта и т. д. – собирается простая репрезентация лица. В-третьих, нейроны возбуждаются в ответ на изображение лица, расположенное в определенном месте их рецептивного поля. В-четвертых, некоторые клетки достигают частичной инвариантности к положению лица в пространстве и к углу зрения. В-пятых, нейроны в самом переднем участке распознавания лиц достигают почти полной инвариантности в отношении указанных аспектов. Наконец, у людей близлежащая область – одна из мишеней передней части височной коры – содержит клетки, реагирующие только на небольшой ряд знакомых лиц. Таким образом, складывается впечатление, что главная цель всех этих участков распознавания лиц в том, чтобы поэтапно формировать репрезентации индивидуальных идентичностей – людей или объектов.
Наверняка вы обратили внимание на степень расплывчатости при описании этой гипотетической нейронной сети. Дело в том, что мы далеки от механистического понимания высших этапов обработки зрительной информации как основанных только на специфических нейронах и синапсах. На самом деле во многих отношениях очевидно, что мозг не может полагаться на простые перцептронообразные нейронные сети, используемые компьютерами для распознавания лиц и управления автомобилями. Забегая вперед, скажу, что, в отличие от большинства искусственных нейронных сетей, опирающихся на контролируемое обучение, мозг обучается без учителя. Я хотел здесь не столько описать конкретную форму нейронной сети, сколько подчеркнуть общий принцип, который заключается в том, что распознавание объектов основано на мультинейронных ансамблях, сформированных посредством постепенной модификации и усиления синаптических связей, как это и предполагал Хебб.
Напомню, что лица – не единственные объекты, распознаваемые в височной коре. Другие участки специализируются на широком разнообразии других типов объектов, связанных визуально или концептуально. Хороший пример – клетки, реагирующие на изображения инструментов, причем не на конкретные инструменты, а на инструменты как категорию (молотки, пилы, плоскогубцы). Мы только начинаем делать первые шаги к расшифровке необычной логической схемы височной доли.
Часть III
До самого горизонта и дальше
Как вы могли заметить, по ходу этой книги мы постепенно перемещались от признанных научных фактов к все менее и менее изученным феноменам. Далее мы собираемся вступить и вовсе на зыбкую почву предположений и догадок. Но зачем нам предаваться этим опасным «спекуляциям», которых серьезные нейробиологи чураются как огня? Если мы этого не сделаем, вся рассказанная выше длинная подробная история останется оборванной на самом интересном месте: что происходит дальше в цепочке зрительного восприятия. К сожалению, никто не знает этого наверняка, но в третьей части книги я постараюсь обрисовать вам в общих чертах то, что нам известно об этой малоизведанной территории, где восприятие сливается с мышлением.
12 | Почему эволюция так любит нейронные сети
Выживают не самые сильные виды и не самые умные, а те, которые лучше других приспосабливаются к изменениям.
ЧАРЛЬЗ ДАРВИН
Разумно предположить, что за несколько миллионов лет эволюции природа могла сотворить все, что хотела. Почему же она выбрала нейронные сети? Если кратко: природа решила, что гораздо проще, экономичнее и эффективнее создать модифицируемый синапс, чем перестраивать геном – генетическую схему зрительной системы – для каждого живого вида. Способная к обучению нейронная система мозга, по сути, представляет собой универсальный механизм, который может гибко адаптироваться к специфике различных визуальных сред (ближнее зрение в лесу, дальнее зрение на равнине) и одновременно распознавать ваше любимое чадо среди других детей на детской площадке.
Первое ключевое преимущество такого устройства касается развития мозга. Представьте себе альтернативу: мозг, состоящий из фиксированных связей, – машина для распознавания лиц, похожая на красивый швейцарский хронометр XVIII в. со множеством мелких латунных деталей, где каждое знакомое лицо кодируется особой комбинацией колесиков и шестеренок. Но тут возникают серьезные загвоздки: во-первых, этих комбинаций вряд ли бы хватило на всех людей, которых человек узнает в течение жизни. Во-вторых, чтобы машина узнала этих людей, они должны быть запрограммированы в ней заранее. В случае с человеческой машиной это означает, что генеральный строительный план – генетический код, превращающий оплодотворенную яйцеклетку в человека, – должен содержать информацию о каждом потенциальном знакомом лице. Но как природа может знать об этом наперед?
Это также нереалистично с точки зрения объема генетического кода. Первичная зрительная кора содержит примерно 14 млн нейронов, которые связаны со вторичной зрительной корой (V2), содержащей около 10 млн нейронов. Если бы система имела фиксированные связи – то есть если бы аксон каждого нейрона V1 имел строго заданную мишень в V2, – процесс наведения аксонов требовал бы огромного количества дифференцированных молекулярных сигналов, каждый из которых пришлось бы кодировать определенной последовательностью ДНК. Предполагать, что такой уровень специфичности действительно может существовать, просто нелепо. Даже несмотря на умные способы архивирования, информационная емкость нашего генома с его жалкими 20 000 генов на порядки меньше тех объемов информации, которые потребовались бы для кодирования одной только системы зрительного восприятия.
Поэтому вместо того, чтобы пытаться изначально специфицировать все возможные связи, природа использует двойную стратегию. Сначала в действие вступают генетически запрограммированные правила для механизма аксонального наведения (молекулярные пути, диффундирующие сигналы), благодаря которым аксоны нейронов-источников соединяются с нейронами-мишенями в заданной области мозга. Именно поэтому участки распознавания лиц в височной доле приматов находятся примерно в одном и том же месте. И именно благодаря этому топография каждой зрительной зоны отражает пространственную карту сетчатки. Но этот механизм работает только на грубом уровне. Поэтому после того, как была сформирована грубая схема связей, в действие вступают правила машинного обучения, которые уточняют эти связи и кодируют восприятие конкретных объектов. Другими словами, молекулярные механизмы, если воспользоваться терминологией Ливингстон, создают «протоучастки» распознавания лиц, расположение которых жестко запрограммировано в генетическом коде, тогда как их окончательная избирательность формируется синаптической пластичностью.
Другое ключевое преимущество состоит в том, что нейронные сети эффективно решают проблему запоминания и распознавания визуальных объектов, с которыми человек сталкивается на протяжении жизни и которые могут представать перед ним в самом разном виде, на разных расстояниях, под разными углами. В нейронной сети каждый нейрон может участвовать в нескольких распознающих схемах в зависимости от того, как его вывод обрабатывается следующим слоем. Если в нейронной сети много слоев и каждый слой содержит, скажем, несколько десятков тысяч элементов, это дает нам поистине астрономическое число возможных комбинаций – достаточное для того, чтобы запомнить лицо любимой бабушки во всех возможных ракурсах и чтобы узнать своего ребенка, будь он прыгающим, бегающим и даже чумазым, в толпе других детей на детской площадке.
В самом фундаментальном смысле главное преимущество нейронных сетей состоит в том, что они позволяют зрительной системе мозга адаптироваться к естественной среде обитания данного живого вида. Нейронная сеть, отвечающая за визуальное распознавание, учится распознавать именно те визуальные объекты, которые присутствуют в конкретном окружающем мире этого вида. Это могут быть очень простые объекты, такие как ориентированные края, которые являются важнейшим признаком практически во всех визуальных сценах в жизни млекопитающего. Или более сложные, например, лица, которые играют важную роль в жизни социальных животных, таких как приматы, включая нас с вами. Как показывают эксперименты, если подопытная обезьяна с детства не видит лиц, ее протоструктуры распознавания лиц учатся распознавать руки. Другими словами, эти нейронные системы развиваются посредством обучения.
* * *
Я не могу удержаться от того, чтобы еще раз не подчеркнуть элегантность, экономичность и простоту естественного дизайна сенсорных систем мозга. Мы вновь и вновь убеждаемся в удивительной продуманности лежащего в их основе организационного принципа: зрительная система настраивается на статистически значимые закономерности естественного мира – на те составляющие визуального ввода, которые наиболее важны для выживания данного живого вида. В случае некоторых ключевых компонентов, которые всегда присутствуют в визуальном мире этого вида, эволюция сочла необходимым потратить драгоценные гены, чтобы запрограммировать сетчатку на восприятие этих аспектов. Хороший пример – «вшитая» функция обнаружения контраста (краев), которая реализуется при помощи молекулярных инструкций, эволюционировавших на протяжении многих миллионов лет, от эпохи, когда обитали мечехвосты, до времени появления людей[34].
Но если латеральное торможение – достаточно простой механизм, который легко биологически запрограммировать до рождения, то восприятие сложных объектов требует слишком большого количества настроек, чтобы их можно было закодировать в нашем ограниченном по размеру геноме. У нейронных сетей нет такого ограничения. Согласно правилу Хебба, мозг может адаптироваться к любым существующим закономерностям более высокого порядка. Глаза, носы, волосы, подбородки появляются в нашем визуальном мире не случайным образом – обычно они появляются вместе, что является хорошей предпосылкой для перцептивного обучения.
Вот еще один пример. Посмотрите на рисунок. И представьте себе мир будущего, в котором некоторые часто встречающиеся «лица», например, инопланетян выглядят примерно так. В этом случае наши нейронные сети в височной доле будут обучаться и на этих примерах тоже и, вполне вероятно, станут распознавать эти странные объекты как лица.

Но пока нейронные сети в височной доле землян настроены только на человеческие лица. Даже если существует гипотетическая возможность встречи с инопланетянами, нашему мозгу не имеет смысла заранее выделять нейроны под их узнавание. Наша зрительная система не занимается распознаванием объектов «просто так». Она тратит свои вычислительные ресурсы только на те объекты, которые она видит регулярно и которые критически важны для нашего выживания.
Такой взгляд на зрение носит прежде всего прагматический характер: он обращается к чисто практической проблеме эффективного использования нейронного оборудования. Но в нем есть и некая возвышенная утонченность, поскольку это говорит о том, что мозг человека или животного теснейшим образом, на уровне синаптических связей между нейронами, связан с природным миром, в котором он существует. Благодаря перцептивному обучению наш визуальный мозг содержит встроенную копию, дистиллят окружающего нас мира.
13 | Прогресс есть, тайны остаются
Если ты не знаешь, куда идешь,
то можешь никуда и не прийти.
ЙОГИ БЕРРА
Распознающие лица нейроны в передней височной коре, по сути, решают классическую проблему распознавания объектов, обозначенную нами в главе 1: они реагируют на лицо независимо от его освещенности, ракурса и положения на сетчатке. Но что дальше? Насколько мы продвинулись в нашем понимании того, что большинство людей считают восприятием?
По правде говоря, не так уж и далеко. Наши знания заканчиваются на группе нейронов в височной доле, которые сообщают нам о том, что в нашем поле зрения находится какой-либо конкретный объект. Довольно скудный перцептивный мир, согласитесь. А наши искусственные модели, видящие компьютеры, способны выполнять только очень узкие задачи, причем весьма неуклюжим и неэффективным образом в сравнении с мозгом. Давайте еще раз вернемся к искусственному интеллекту и рассмотрим важнейшую фундаментальную проблему, которую он, несмотря на свои впечатляющие успехи, пока так и не сумел решить.
ОБУЧЕНИЕ С УЧИТЕЛЕМ И БЕЗ УЧИТЕЛЯ
Мы почти ежедневно слышим об искусственном интеллекте и различных перцептивных задачах, которые он способен выполнять при помощи машинного обучения: беспилотные автомобили, распознавание лиц и многое другое. Все это сопровождается паническими пророчествами, что, дескать, умные машины однажды станут умнее людей, выйдут из-под контроля и захватят мир. Но большинство таких прорицателей не замечают в комнате огромного слона: машинное обучение действительно способно делать некоторые замечательные вещи, но, как это мучительно осознают специалисты в области ИИ, сегодня даже самые умные машины куда глупее четырехлетнего ребенка.
Причина в том, что даже наиболее продвинутые алгоритмы ИИ требуют для своего обучения колоссальных объемов данных и сверхмощных компьютеров. Четырехлетний ребенок учится сам, причем зачастую ему бывает достаточно всего нескольких примеров. В данном случае речь идет не о навыках, знаниях, правилах и пр., которым мы целенаправленно и старательно учим детей, а о базовых механизмах восприятия, которым их маленький мозг учится совершено самостоятельно. В отличие от многослойного перцептрона с его механизмом обратного распространения и т. п., которому требуется несметное количество примеров и помощь учителя, чтобы научиться распознавать одно лицо, моему маленькому внуку хватило нескольких теплых объятий, чтобы при виде меня кричать: «Деда!»
Исследователи ИИ различают два типа обучения: контролируемое (с учителем) и неконтролируемое (без учителя). Как вы помните, наш простой канонический перцептрон обучается с помощью учителя. Как и программа распознавания речи Apple, говорящий компьютер Сейновски и системы распознавания лиц, пугающие защитников конфиденциальности. В последнем случае в роли учителя выступают огромные каталоги лиц с индивидуальными идентификаторами. Компьютеры могут учиться таким способом, потому что они очень быстрые – на самом деле своими недавними достижениями ИИ в значительной мере обязан появлению в последние пять лет гигантских наборов обучающих данных и мощных специализированных компьютеров. Мозг со своими медленно работающими нейронами никогда не сможет конкурировать с ИИ на этом поле.
Но во всем прочем мозг превосходит ИИ. Оригинальная концепция Хебба объясняет, как мозг учится без учителя: в ответ на естественные закономерности видимого мира, такие как края, мозг создает чувствительные к краям нейроны и связывает их в сеть, клеточные ансамбли. В настоящее время исследователи ИИ стараются создать машины, способные к такому же неконтролируемому обучению, как мозг[35].
Вскоре мы вернемся к живому мозгу, но сначала давайте рассмотрим еще один способ машинного обучения, использующий немного другой принцип. Создатели польстили своим алгоритмам, назвав их «обучение с подкреплением»: это название отсылает нас к механизму обучения, который впервые был описан великим русским физиологом Павловым и глубоко изучен его последователями, в частности, Б.Ф. Скиннером из Гарвардского университета. Суть обучения с подкреплением проста: когда и если определенное поведение вознаграждается, оно начинает повторяться. Повторение поведения приводит к усилению синаптических связей в соответствующей нейронной сети. В принципе, подкрепление является формой обратного распространения, а обучение с подкреплением отличается от контролируемого обучения лишь тем, что компьютер сам генерирует себе учителя.
Компьютеры тоже можно вознаграждать. Например, компьютеру задают некую задачу. Он делает первую попытку, как правило, неудачную. Но, если попытка хотя бы немного приближает его к поставленной цели, его вознаграждают. Конечно, его не балуют каким-то лакомством, а вместо этого говорят: «Окей, увеличь синаптические веса того, что ты только что сделал». Затем компьютер делает новую попытку, на этот раз с новыми весами. Так, попытка за попыткой, компьютер уточняет свои веса, пока в конце концов не научится выполнять поставленную задачу.
Хотя этот процесс кажется примитивным, основанные на обучении с подкреплением алгоритмы уже овладели поразительно сложными умениями: они научились играть в шахматы и даже в го (игру, которая по сложности превосходит шахматы). Эти компьютеры не только играют на сверхчеловеческом уровне (они уже доказали свою способность победить любого человека), но и научились играть совершенно самостоятельно. В журнале Nature незадолго до Рождества 2018 г. рассказывалось об одной из таких программ под названием AlphaZero. Программе задавались только правила игры – определение игрового поля, правила ходов для каждой фигуры и т. д., после чего она начинала играть в шахматы или го сама с собой. Звучит немного странно, но хитрость заключалась в том, что каждое из двух «я» не знало, что думает другое «я», – оно видело только сделанный ход. Вместо внешнего учителя использовались внутренние правила, которые говорили программе, был ли данный ход удачным или нет и кто выиграл игру. AlphaZero понадобилось всего четыре часа, чтобы научиться играть в шахматы не хуже гроссмейстеров мирового уровня.
Эта удивительная способность применима не только к настольным играм, но к множеству других задач. В одной из своих лекций Дэвид Силвер из команды Google AI показал видео, на котором AlphaZero дистанционно управляет игрушечным вертолетом, заставляя машинку выполнять фигуры высшего пилотажа. Наблюдая за акробатическими трюками вертолета в воздухе, вы готовы поверить всем мрачным прорицаниям по поводу ИИ.
Но может ли AlphaZero соперничать в уме с моим внуком? Нет, даже отдаленно (если только мой внук не решит сразиться с ним в шахматы). Компьютеру нужна очень узко поставленная задача. Его мозг намного больше, чем мозг моего внука, и для работы ему требуется куда больше энергии, чем тарелка молочной каши. По оценке Кэтрин Ву в журнале Smithsonian, оборудование AlphaZero потребляет около одного миллиона ватт; мозг моего внука – меньше 20 ватт. Зачем же тогда я вообще затронул тему нейронных сетей и обучения с подкреплением (AlphaZero также построена на внутренних нейронных сетях)? Дело в том, что они служат так называемыми доказательствами принципа действия – доказательствами того, что данный тип логики может генерировать функциональность, хотя бы отдаленно приближающуюся к функциональности живого мозга.
Может ли человеческий мозг выполнять те же манипуляции, что и глубокие нейронные сети или AlphaZero? Разумеется, может, хотя и гораздо медленнее. Человеческий мозг – это компьютер, созданный за миллионы лет эволюции, которая довела до совершенства и миниатюризировала его синаптические связи. Если что-то может быть сделано громоздкими компьютерными микросхемами, это может быть сделано и мозгом.
Разработчики ИИ прекрасно осознают, что на сегодняшний день мой внук превосходит их компьютеры по всем параметрам, поэтому трудятся в поте лица. Я думаю, что машины в конце концов станут чертовски умными – но что именно они научатся делать, пока мы можем только гадать. Сегодня ведется разработка удивительных способов машинного обучения без учителя. Вопрос только в том, сколько времени это займет и насколько машины смогут сымитировать работу мозга. И, самое главное: сможет ли компьютерное оборудование хотя бы немного приблизиться в экономичности к нейронному? Я без страха смотрю в будущее. На самом деле именно фактор энергопотребления позволяет мне не бояться того, что однажды компьютерный сверхразум захватит мир.
ВИНФРИД ДЕНК И КОННЕКТОМ
Мало кто может сравниться с Винфридом Денком, директором Института нейробиологии Общества Макса Планка в Мартинсриде, Германия, в креативности научного мышления. Вот его секрет: самые плодотворные творческие идеи приходят ему в голову между полуночью и четырьмя часами утра.
Денк – высокий, крепкого телосложения человек с взъерошенной шевелюрой и аккуратной бородкой. Его редко увидишь без улыбки. Поиск в Google выдает всего одну фотографию. На ней Винфрид предельно серьезен, одет в белую рубашку, пиджак и галстук, притом что обычно он предпочитает джинсы и рубашки в стиле кэжуал. Строгий официальный портрет был сделан по случаю присуждения исследователю премии Кавли, одной из самых престижных наград в области нейронаук, – вероятно, учредители премии потребовали, чтобы Денк предстал в образе «настоящего» серьезного ученого.
Эту и другие награды Денк получил за замечательную серию научных разработок, в основном в области физики и оптики. Его первым достижением стало усовершенствование конфокального микроскопа. Конфокальный микроскоп, о котором уже упоминалось выше, представляет собой разновидность светового микроскопа, но с гораздо более высоким разрешением. Конфокальная микроскопия соединяет оптическую микроскопию и компьютерный анализ – в ней нет изображения в традиционном смысле; вместо этого производится точечное сканирование образца, и эти серии отсканированных точек собираются вновь и дают цифровое изображение. Технология быстро завоевала сердца исследователей и в настоящее время стала отраслевым стандартом.
Эту работу Денк проделал, будучи постдоком в Корнеллском университете, под руководством наставника Уотта Уэбба. Конфокальная микроскопия существовала и раньше, но скорее в виде полутеоретических разработок; Винфрид превратил ее в удобный, полезный инструмент для биологов. Следующее изобретение – двухфотонный лазерный микроскоп – он разработал и запатентовал вместе с Дэвидом Танком во время короткого периода работы в знаменитом исследовательском центре Bell Labs, который сегодня, к сожалению, лишился своего статуса мировой фабрики идей и инноваций, пав жертвой интересов большого бизнеса. Двухфотонная микроскопия обладает гораздо большей проникающей способностью, чем конфокальная: она позволяет глубже заглянуть внутрь толстых образцов, с меньшим повреждением тканей.
Затем Денк вернулся в Германию и возглавил Институт медицинских исследований Макса Планка в Гейдельберге. Получив в свое распоряжение значительные ресурсы, он нанял команду инженеров, программистов и техников и расширил масштабы своей научной деятельности, занявшись не одним, а сразу двумя крупными проектами.
В первом Денк объединил силы с младшим сотрудником Томасом Эйлером, чтобы с помощью двухфотонной микроскопии попытаться разрешить полувековую загадку такого свойства сетчатки, как избирательность в отношении направления. Для этой цели Денк и его коллеги выбрали так называемый звездчатый тип амакриновых клеток. Стимулируя сетчатку движущимся раздражителем, они наблюдали за одной из таких клеток через микроскоп. (Это можно было сделать только с помощью двухфотонного микроскопа, поскольку тот использует свет с такой длиной волны, которая почти не возбуждает палочки и колбочки. Излучение обычного светового микроскопа засвечивало бы фоторецепторы, и те бы не реагировали на тестовый стимул.) Благодаря предварительно введенному в сетчатку индикатору активности исследователи могли видеть, когда звездчатая клетка реагировала на тестовый раздражитель. К своему удивлению, они обнаружили, что те амакриновые клетки, которые синаптируют с избирательными к направлению ганглионарными клетками, сами являются избирательными в отношении направления: если не вдаваться в подробности, именно эти звездчатые клетки заставляют ганглионарные клетки быть чувствительными к направлению.
Одновременно Денк занимался разработкой инновационной технологии – на этот раз не микроскопа, а устройства, позволяющего обойти некоторые сопряженные с использованием микроскопов проблемы. Его изобретение может внести поистине огромный вклад в развитие нейронауки.
Но прежде, чем углубиться в эту область, которая получила название коннектомика, давайте поближе познакомимся с modus operandi Денка. Как я уже сказал, он работает по ночам – хороший способ выкроить несколько часов тишины и покоя. Но что он делает в своем кабинете в два часа ночи? Нет, он не тратит это драгоценное время на стандартную профессорскую рутину вроде подготовки к лекциям, рецензирования рукописей и т. п. Что касается руководства институтом, то он поручает всю трудоемкую или нетворческую работу своим сотрудникам. По ночам он читает и размышляет – то, чем большинство из нас занимается лишь урывками, Денк делает обстоятельно и глубоко.
Еще Денк проводит много времени в самолетах, потому что его приглашают выступать с лекциями по всему миру. Не сказать чтобы ему нравилась работа лектора; он принимает приглашения, потому что они дают ему возможность пообщаться с нужными людьми. Винфрид – гениальный, как сейчас модно говорить, «шмузер», у которого есть свой метод. Он находит людей, изучающих важные для него научные проблемы, и старается вывернуть их ум наизнанку. Я не раз слышал, как, прознав о новом интересном исследовании, он говорил, что поедет пообщаться с его автором, даже если это был совершенно незнакомый ему человек, проживавший где-нибудь на западном побережье США или в Китае.
Поскольку именно я подсказал Денку заняться нейронами сетчатки с избирательностью в отношении направления, он на протяжении нескольких лет удостаивал меня регулярными визитами. Завершив свое исследование и опубликовав большую статью в журнале Nature, он снова явился ко мне и сказал: «Я разгадал твою проблему. Что будем делать дальше?» К сожалению, на тот момент мне нечего было ему предложить – по крайней мере ни одна из имевшихся в моем поле зрения проблем не могла сравниться в важности с проблемой избирательности к направлению. После этого его визиты прекратились. Не примите это за упрек в его адрес! Визиты Винфрида невероятно полезны нам обоим. Я готов с ним «шмузить» – поболтать на научные темы – в любое время.
В год он получает несколько сотен приглашений: если бы он принимал их все, у него бы не осталось времени ни на что другое. Выступления Винфрида притягательны тем, что он к ним почти не готовится. Он бродит по сцене, периодически утыкается взглядом в пол, иногда забывает, каким будет следующий слайд, и, кажется, говорит все, что приходит в голову. Нет, он умеет формулировать свои мысли с кристальной четкостью – и делает это в письменных работах и профессиональном общении. Просто он не желает тратить время на подготовку столь модных сегодня презентаций. Винфриду всегда есть что сказать, но он предпочитает давать слушателям больше свободы. Я уважаю его за это.
Теперь о коннектоме (connectome). Окончание -ome означает «совокупность», в данном случае – это совокупность всех связей. Таким образом, коннектом есть полное описание всей структуры нейрональных связей в нервной системе организма. Составление коннектома человека – процесс, захватывающий дух своей дерзостью, объемом работы и перспективами, и он вряд ли будет завершен при нашей с вами жизни. Но Винфрид изобрел способ, как это можно сделать, а неодолимая сила научного прогресса позаботится обо всем остальном.
Как определить, связаны ли два нейрона? Для этого нужно найти синаптический контакт между ними, что очень непросто, учитывая крошечные размеры синапсов – примерно от 0,5 до 1,0 мкм. При таких размерах приходится использовать электронную микроскопию, и в этом вся загвоздка. Как вы уже знаете, традиционная электронная микроскопия требует разрезания тканевого образца на ультратонкие срезы толщиной примерно 50 нанометров. Это означает, что нужны сотни и даже тысячи последовательных срезов, чтобы охватить всего лишь один синапс, не говоря уже о расстоянии между двумя клетками. Используя традиционные методы, ни один человек не в состоянии нарезать достаточно срезов, чтобы охватить это расстояние, и, даже если это будет сделано, как затем правильно выровнять эти срезы относительно друг друга?
Денк придумал способ решить эти две проблемы одним махом. Разработанный им метод называется сканированием лицевой поверхности блока. В нем так же используется растровая электронная микроскопия, но, в отличие от традиционного подхода, нет необходимости разрезать образец ткани на тысячи тончайших частей: вы сканируете поверхностный слой тканевого блока, после чего срезаете (и выбрасываете) этот слой. В результате перед вами остается новая лицевая поверхность, которую вы снова сканируете электронным микроскопом. Весь процесс резки автоматизирован. Это также решает проблему совмещения изображений: поскольку тканевой блок остается стационарным, при каждом очередном срезе изображение практически не сдвигается.
Таким образом, метод Денка позволяет получить длинный ряд изображений, представляющих серийные срезы тканевого образца с высочайшим разрешением электронной микроскопии. Разумеется, после этого необходимо преодолеть еще множество технических трудностей, но главное, что это дает нам легко выравниваемые изображения, на которых можно проследить нейронные связи – на скромных пока расстояниях, которые, как мы надеемся, значительно увеличатся в будущем. Каждый срез содержит сотни и даже тысячи нейронов, поэтому их прослеживание – колоссально трудоемкая работа (сложная даже для компьютеров). Но в принципе, если мы проделаем эту черновую работу, в конечном итоге мы получим полную карту всех нейрональных связей.
Почему это так важно для нейробиологии? Потому что по большому счету это все, что у нас есть. Мозг – машина связей; если мы будем знать все его связи, мы сможем значительно продвинуться на пути к пониманию того, как работает эта сложнейшая биологическая машина. Винфрид опробовал данный метод на сетчатке глаза и успешно показал, как синаптические связи определяют такое свойство сетчатки, как избирательность к направлению, – причем показал это с такой степенью конкретности и надежности, которая ранее была недостижима при исследовании нейронных сетей.
С момента изобретения метод Денка был модифицирован и усовершенствован, и сегодня ряд лабораторий использует его в разных вариациях для решения различных задач. Но главной заслугой Денка являлся концептуальный прорыв: осознание возможности осуществить полное картографирование и анализ всех связей, которые лежат в основе работы головного мозга. Разумеется, нам еще предстоит решить массу технических проблем, например, придумать надежные способы отслеживания дальних связей – сначала между соседними областями мозга (скажем, между зонами V1 и V2), а затем и между более удаленными друг от друга. И даже тогда останутся важные вопросы: какие нейромедиаторы используются в тех или иных синапсах? Какие сообщения они несут? Но все это обязательно произойдет. Я не сомневаюсь: в конце концов – в очень долгосрочной перспективе – коннектом станет фундаментом, который ляжет в основу нашего понимания любых сложных нейронных сетей.
Что Винфрид Денк будет делать дальше? Какие новые и неожиданные идеи придут ему в голову в ночные часы глубоких размышлений? Кто знает, но, судя по прошлым достижениям, это будет чем-то захватывающим.
НЕЙРОННЫЕ СЕТИ ПОД МИКРОСКОПОМ
Область машинного обучения – это поприще гиков-белоручек, на три четверти специалистов по информатике и на четверть – нейробиологов. Исследованием интеллекта занимаются люди другой породы – те, кто не прочь заняться черной работой и хочет знать наверняка, на основе осязаемых физических фактов, как работает мозг.
Первый вопрос, который обычно приходит в голову экспериментальному исследователю мозга вроде меня: «Как, черт возьми, можно изучать эти нервные сети, которые состоят из десятков тысяч нейронов, разбросанных по всему мозгу? Регистрировать активность сразу нескольких тысяч нейронов? И, будь это даже возможно, как потом переработать все эти данные?» Десять лет назад такое действительно казалось нереальным, но сейчас мы начинаем делать первые шаги в этом направлении.
Как часто бывает, прогресс стал возможен благодаря объединению достижений из нескольких очень разных областей. Во-первых, двухфотонная лазерная микроскопия не только обеспечила более высокое разрешение по сравнению с традиционным микроскопом, но и благодаря своей проникающей способности дала нам возможность заглянуть вглубь тканевых образцов; в случае с мозгом это означает, что мы можем увидеть несколько слоев коры. Как вы уже знаете, первая версия такого микроскопа была разработана Винфридом Денком, одним из самых креативных умов в научном мире и (я надеюсь) будущим лауреатом Нобелевской премии. Некий исследователь сравнил разницу между традиционной и двухфотонной микроскопией с просмотром цветного телевизора в ярко освещенной и темной комнате. Еще одно преимущество двухфотонной микроскопии в том, что она позволяет наблюдать клетки, не повреждая их (что является проблемой при использовании традиционного конфокального микроскопа).
Во-вторых, благодаря генной инженерии мы получили возможность встраивать в нейроны мозга белковую молекулу, которая мерцает (говоря научным языком, меняет свою флуоресценцию) при активизации нейрона. Если смотреть через конфокальный микроскоп на образец генно-модифицированной нервной ткани, в которую встроена такая молекула, можно в режиме реального времени наблюдать за активностью отдельных нейронов.
Третьим прорывом мы обязаны биологам, изучающим поведение насекомых. Представьте, что вам нужно отследить траекторию движения жука, чтобы понять, какие сигналы заставляют его поворачивать налево, а какие – направо. Один из способов – поручить своему постдоку снять на видео бегущего жука и затем детально описать его маршрут. Но экспериментаторы придумали, как автоматизировать этот процесс. На панцирь жука приклеивается стационарная платформа так, чтобы его лапы болтались в воздухе. Под жуком помещается легкий шарик наподобие мячика для пинг-понга, который подвешен в специальном держателе, практически не создающем трения, так что бегущий жук легко вращает его своими лапами. Компьютер измеряет все повороты мяча, а ваш постдок в это время может заняться чем-то более увлекательным.
Четвертым достижением, которое сегодня мы воспринимаем как данность, стала дешевая вычислительная мощь. Когда двухфотонный микроскоп выдает вам изображения нескольких тысяч клеток, каждая из которых мерцает в соответствии со своим индивидуальным паттерном активности, вы получаете не просто большие, а астрономически большие данные. Без современных вычислительных мощностей мы бы не извлекли из этих гор сырой информации и крупицы полезных знаний.
И, наконец, потребовался талантливый, настойчивый и дерзкий нейробиолог по имени Дэвид Танк из Принстонского университета, один из создателей двухфотонной микроскопии, который соединил вместе все вышеописанные достижения и добавил от себя несколько замечательных новшеств.
«Почему бы нам не нацелиться на звезды? – подумал Танк. – Что, если нам попытаться разглядеть сразу тысячи нейронов у животного, которое находится в сознании, видит разные зрительные стимулы и думает о них?» Вместе с коллегами Танк придумал способ, как зафиксировать мышь в ограничивающей рамке наподобие той, которую инсектологи используют для жуков. Мышь не особенно возражает, потому что ее кормят лакомствами. Как и жук, она стоит на свободно вращающемся шаре. Это генно-модифицированная мышь – в ее корковые нейроны методом генной инженерии встроены молекулы – индикаторы активности. Вокруг себя мышь видит искусственный мир, созданный с помощью технологии виртуальной реальности. И да, у нее в черепе проделано отверстие, через которое Танк и его коллеги с помощью конфокального микроскопа могут наблюдать за активностью корковых нейронов. Исследователи обучают мышь различным задачам – в частности, бегу по виртуальному лабиринту – и следят за тем, как ведут себя ее нейронные сети.
Хотя этот метод еще довольно сырой, он работает. И первое фундаментальное открытие состоит в том, что нейронные сети могут вести себя поразительно стабильным образом: например, в большинстве случаев в ответ на определенный стимул вспыхивает одна и та же группа первичных сенсорных нейронов. Это удивило исследователей, поскольку они предполагали, что нейроны будут мерцать в соответствии с собственной, непонятной нам логикой. Действительно, нейроны в корковых областях, отвечающих за сенсомоторную интеграцию, демонстрируют более гибкие, еще полностью не расшифрованные паттерны активности, иногда устойчивые, иногда меняющиеся. Это вполне объяснимо, так как эти области связывают сенсорный вход и поведенческий ответ на него, который может варьироваться в зависимости от ситуации. Но все это уже концептуальные проблемы, а не технические преграды.
Здесь я прерву свой рассказ. Ученики Танка внедряют этот метод в своих нейробиологических лабораториях; мыши бегают по виртуальным лабиринтам; компьютеры регистрируют паттерны активности их нейронов. От открывающихся перспектив у нейробиологов вскипают мозги. Сможем ли мы увидеть, как образуются клеточные ансамбли? Где в головном мозге хранятся воспоминания – в каком-то одном месте или же они мигрируют? Что делают нейроны, когда мышь спит, когда она чего-то хочет или когда видит другую мышь? Теперь у нас есть инструмент, и все, что нам нужно, – это экспериментировать и еще раз экспериментировать. Оставайтесь на связи: пока вы читаете эту книгу, эксперименты продолжаются!
14 | Зрение и сознание
Где-то что-то невероятное ждет своего познания.
БЛЕЗ ПАСКАЛЬ
Мне неизвестно, что может уготовить нам будущее, но, что бы там ни было, я иду навстречу этому смеясь.
ГЕРМАН МЕЛВИЛЛ
За всем тем, что сказано было здесь о восприятии, маячит один ключевой вопрос: кто наблюдатель? И где он «находится» в головном мозге? Не только изучать восприятие, но и размышлять о его глубинной природе – естественно для нас, ученых и обычных людей. Изучая восприятие, мы выступаем в роли внешнего наблюдателя: мы находимся за пределами мозга (или компьютера), наблюдаем за нервными импульсами (или электронами), бегущими туда-обратно по нейронным сетям, и пытаемся выяснить, как все это отражает внешний мир. И это вполне выполнимая задача – понять, как нейроны мозга представляют реальный физический мир, чтобы помочь нам не попасться в лапы хищника или пройтись по городской улице. Наш мозг составляет точную карту визуальной реальности, которая позволяет нам брать руками нужные предметы или объезжать препятствия, несясь с горы на лыжах. Но как насчет внутреннего наблюдателя? Неужели в нашей голове сидит некий маленький человечек, «гомункулус», который, как зритель в театре, созерцает все данные чувственного опыта?[36]
Пожалуй, самое труднообъяснимое во всем том, что я рассказал вам здесь о зрении, – так называемая проблема связывания. Каким образом мозг связывает все эти разнообразные и разрозненные визуальные входные сигналы, чтобы создать целостное восприятие – например, красного автомобиля, движущегося слева направо?
Разложение визуального ввода на параллельные потоки информации является фундаментальным принципом работы зрительной системы. Давайте еще раз вернемся к сетчатке. Первое, что делает сетчатка, – улавливает свет и сжимает широкий диапазон яркости до узкого рабочего диапазона. Второе – значительная часть сетчатки занимается фрагментацией визуального изображения, разбиением его на отдельные сигналы, представляющие движение, цвет, края и т. д. Это действительно разные сигналы: чувствительная к направлению движения ганглионарная клетка ничего не сообщает мозгу о цвете движущегося объекта, а кодирующие цвет клетки ничего не говорят о наличии или отсутствии краев.
Эти раздельные информационные потоки поступают в разные отделы мозга. Большинство нейронов сетчатки отправляют свои сигналы в латеральное коленчатое тело, откуда те поступают в зрительную кору головного мозга. Но некоторые клетки отклоняются от этой модели и направляют свои аксоны в другие функциональные зоны. Так, клетки, отвечающие за восприятие абсолютной яркости, направляют свои сигналы в центры мозга, регулирующие цикл сна – бодрствования. Другая группа клеток обеспечивает специализированный визуальный ввод для стабилизации глаз. Но все потоки сигналов нигде не сводятся вместе анатомически – во всяком случае они обрабатываются раздельно простыми и сложными клетками зрительной коры, детекторами сложных движений средней височной коры и т. д. Как они снова «связываются» вместе в единое целостное восприятие?
Предложенный нейробиологом Анной Трейсман ответ основан на том факте, что все эти начальные области зрительной системы мозга, судя по всему, содержат топографическую карту сетчатки: такая карта была обнаружена в зрительных зонах V1 и V2, в средней височной области и в более высоких областях височной коры. Что, если эти топографические карты связаны таким образом, что, например, клетки, представляющие цвет, точка в точку совпадают с простыми или сложными клетками в областях V1, V2, V3, V4 и в участках распознавания лиц в височной коре? Эксперименты показали, что такие связи действительно существуют.

На рисунке справа – изображение, которое может создавать группа нейронов в зоне V1, специализирующаяся на выделении границ, и оно немного напоминает схематическое HOG-изображение. Среднее изображение может быть выходом нейронов в сетчатке или ЛКТ, которые специализируются только на усилении краев. Изображение слева дополнено репрезентацией цвета, в данном случае оттенков серого, за что отвечает, в частности, определенный тип клеток сетчатки с устойчивым ответом. Если три данных изображения представлены в разных отделах мозга в виде отдельных топографических карт, мозг может на каком-то этапе накладывать эти карты друг на друга и получать единое изображение, как на рисунке слева.
Вольф Зингер, Кристоф фон дер Мальсбург и другие немецкие коллеги выдвинули иную гипотезу, предположив, что целостное изображение объекта может формироваться благодаря синхронизированному срабатыванию нейронов, создающих отдельные репрезентации. В ответ на репрезентацию выделенных краев возбуждаются нейроны, отвечающие за репрезентацию объекта, а также за цветовую репрезентацию. Их синхронное срабатывание говорит мозгу, что все эти три репрезентации относятся к одному объекту. В настоящее время существование этого механизма пытаются подтвердить экспериментальным путем.
* * *
За всем этим стоит еще более фундаментальная проблема, издавна интересующая людей: что такое наше сознание? Где именно оно находится? Где обитает наше «я»? Большинству из нас кажется, что наша сознательная сущность сосредоточена в голове, в области, расположенной непосредственно за глазами. Не может ли это быть тем самым центром, который решает вышеупомянутую проблему связывания – который собирает все фрагментированные сигналы, поступающие от сетчатки и других отделов мозга, и с поразительной точностью соединяет их, получая целостную репрезентацию внешнего мира? Но как обычный комок мягкой мозговой ткани может создавать мое уникальное «я», центр моей личной вселенной? Это касается не только меня: у каждого из нас есть субъективное ощущение «я существую». Но что такое это «я» и где именно оно существует? Меня не удовлетворяет ни один из предложенных до сих пор ответов.
Возьмем, например, знаменитый тест Тьюринга, якобы направленный на тестирование машинного разума. Вы создаете компьютер, обучаете его имитировать человеческое мышление, после чего предлагаете постороннему наблюдателю поговорить с вашим компьютером (не видя «собеседника») и определить, с кем он говорит – с живым человеком или машиной. Если посторонний наблюдатель не может отличить машину от живого человека из плоти и крови, утверждал Тьюринг, значит, этот компьютер можно считать мыслящим. Честно говоря, тест Тьюринга всегда напоминал мне голого короля, которому явно чего-то не хватает. Тьюринг, безусловно, был блестящим математиком, но этот тест не имеет ничего общего с математикой. Что конкретно дает основания полагать, будто способность компьютера к совершенной имитации делает его мыслящим? Он по-прежнему остается не чем иным, как куском кремния.
Хебб считал, что наша личность – наше «я» – представляет собой всю совокупность связанных воедино клеточных ансамблей, от самых простых, перцептивных, до гораздо более сложных, представляющих мышление, память, эмоции и мириады других вещей, которые делают нас людьми. Эта концепция по крайней мере хороша своей конкретностью, и мне нравится идея о том, что наше сознание содержит в себе весь наш прошлый опыт. Да, с учетом всего того, что сегодня нам известно о специализации мозга, модель Хебба требует некоторой модернизации – его глобальный клеточный ансамбль должен включать небольшие специализированные регионы, соответствующие конкретным функциональным областям в коре мозга. Однако сама концепция Хебба, что весь мозг может работать как набор нейронных сетей, остается верной.
Итак, мы проследили путь зрительной информации до того места в височной коре, где находятся нейроны, реагирующие на конкретные лица (или другие классы объектов). Логично предположить, что эти распознающие лица нейроны встроены в клеточные ансамбли, потому что одиночный нейрон сам по себе вряд ли способен справиться с такой сложной перцептивной задачей. Но что происходит со зрительной информацией дальше?
Дальше она попадает в следующие клеточные ансамбли. Хебб считал, что между восприятием, мыслью и действием – под последним понимаются нейронные команды, которые заставляют наши мышцы двигаться определенным образом, – нет четко выраженных границ. Все это представлено взаимосвязанными и взаимопроникающими нейронными сетями. На рисунке ниже проиллюстрирована эта концепция (Хебб, вероятно, уже устал вертеться в гробу, поэтому назовем ее «нео-Хеббовской»).

Точки на схеме означают нейроны, линии между ними – аксональные связи. Обратите внимание, что отдельные функциональные области частично переплетаются между собой: перцептивные клеточные ансамбли взаимопроникают с когнитивными; когнитивные ансамбли – с моторными. Слева показаны начальные отделы зрительной системы коры V1–V4, где осуществляется предварительный анализ визуального изображения. В первичной зрительной коре V1 появляются чувствительные к ориентированным краям нейроны. Также есть свидетельства того, что здесь присутствуют нейроны и с более сложной избирательной чувствительностью, которые, в частности, помогают «сегментировать» изображение, разбивать визуальную композицию на дискретные объекты. Клетки в V2 также реагируют на ориентированные края, но с некоторой пространственной инвариантностью. Клеткам в V3 и V4 свойственна весьма разнообразная селективность, так что даже самые опытные экспериментаторы не сумели определить доминирующую специализацию этих двух областей. Если говорить обобщенно, клетки в зрительных зонах коры V1–V4 чувствительны к конкретным составляющим визуального ввода, таким как края, углы, кривизна, цвет, а также почти наверняка к множеству других элементов и признаков изображения.
Проводящие пути в нейронной сети представляют собой клеточные ансамбли, образованные в результате перцептивного обучения по правилу Хебба. Они могут быть короткими, состоящими всего из нескольких нейронов, или большими, пересекающими границы разных функциональных зон. Эти проводящие пути идут в самых разных направлениях, через всю сеть. Так, существуют многочисленные прямые и обратные связи между клетками V1–V4 и зрительными областями височной коры. Обратите внимание, что между клеточными ансамблями начальных зрительных зон V1–V4 и височной коры нет четкой границы. Ансамбль клеток, расположенный в основном в височной коре, может соединяться с одним или несколькими ансамблями в зонах V1–V4. На самом деле именно так выделенные на ранних этапах признаки связываются с репрезентациями более высокого порядка. При активации определенного набора нейронов, отвечающих за выделение определенных первичных признаков, активируются образованные посредством перцептивного обучения связи. Они возбуждают нейроны более высокого уровня, которые отвечают за более общую репрезентацию – в данном случае за репрезентацию морды волка.
Эти клеточные ансамбли, в свою очередь, соединены с ансамблями, которые отвечают за абстрактное мышление, слабо связанное с восприятием. Благодаря такому взаимопроникновению нейронные сети могут возбуждать друг друга. Разумеется, большинство нейронов в «перцептивной» сети связаны преимущественно с такими же перцептивными нейронами, но некоторые из них поддерживают связь с нейронами «мыслительной» сети. Вот почему некоторые перцептивные события могут инициировать мысли – и в действительности даже вызываться мыслями. Иначе говоря, мысль способна вызвать активацию перцептивной нейронной сети, так что мы можем вспомнить или вообразить себе сенсорное событие. Некоторые «мыслительные» клеточные ансамбли соединены с моторными сетями, благодаря чему они могут вызывать их активацию и, как следствие, соответствующие двигательные акты.
Таким образом, клеточные ансамбли, представляющие простые визуальные признаки, соединяются с ансамблями, представляющими репрезентации более высокого порядка, которые, в свою очередь, соединяются с клеточными ансамблями, отвечающими за физические двигательные акты, причем такое соединение всегда происходит в результате перцептивного обучения – усиления синаптических связей в результате одновременной активации сигналами из реального мира в соответствии с правилом Хебба. Вот что я имел в виду, когда говорил, что в концепции Хебба нет четко выраженной границы между восприятием и действием.
Это, конечно же, упрощенная картина, цель которой – продемонстрировать суть концепции Хебба. Как уже отмечалось выше, она требует некоторого уточнения с учетом последних данных, предположительно указывающих на существование очень конкретной функциональной специализации некоторых локальных областей мозга. Однако также необходимо отметить, что исследования с использованием инструментов мультиклеточной визуализации, о которых мы говорили в главе 13 и которые позволяют изучить поведение нейронов в их родных сетях, скорее подтверждают нарисованную здесь картину. Эта картина также больше соответствует реальной анатомии мозга с его непостижимо огромным количеством прямых и обратных связей как между корковыми областями, так и с подкорковыми центрами.
Теперь давайте посмотрим на ту же последовательность сквозь призму современной концепции нейронной сети – более-менее случайной сети связей, состоящей из входного слоя, нескольких скрытых слоев и выходного слоя. Как и в предыдущем примере, это – упрощенная абстракция, призванная проиллюстрировать несколько базовых концептуальных принципов алгоритма.

Как видите, это очень напоминает последовательную схему распознавания лица Авраама Линкольна на с. 102 с тем лишь отличием, что здесь мы вышли за пределы распознавания лиц и включили восприятие, мысли и действия. Перед нами универсальная схема нейронной сети, которая может распознавать не только волка, но и многое другое в зависимости от того, какие синаптические связи были усилены на этапе ее обучения. Как и в концепции Хебба, здесь нет жестких границ между слоями, благодаря чему цепочка синаптических связей может быть изменена на основе опыта от одного конца до другого – посредством обратного распространения или какого-либо другого механизма. Нейронная сеть мозга обучается без учителя, то есть она сама себе учитель. Но как именно это происходит, нам пока неизвестно. Возможно, сеть использует обучение с подкреплением или какой-либо другой метод неконтролируемого обучения из тех, что в настоящее время разрабатываются для искусственных нейронных сетей.
* * *
Итак, вот что я хотел до вас донести: концепция, расплывчатая в отношении большинства конкретных деталей, гласит, что все шаги (от визуального ввода до действия) реализуются через последовательность модифицируемых нервных связей – клеточных ансамблей или слоев нейронной сети. Помимо чисто практических преимуществ – компьютеры уже доказали, что такая сеть способна действовать довольно умно, – чем еще может быть полезна эта концепция? Важнейший момент в том, что в этих моделях нет центра принятия решений, маленького человечка, который принимает сигналы и отправляет команды. Это сеть связей в подлинном смысле.
Но к чему это приводит нас в том, с чего мы начали это обсуждение, – в поиске нейронной идентичности нашего сознания, того «я», которое живет у нас в голове? Предположим на миг, что наш мозг устроен именно так, как показано на двух вышеприведенных схемах. Есть ли в них место сознанию? К сожалению, нет. По сути, это абсолютно бихевиористские модели: ощущение ведет к восприятию, которое ведет к мысли, которая ведет к действию. Все эти этапы не требуют физического разделения; они могут существовать как части, пусть даже как распределенные части, одной нейронной сети. Их вполне можно представить как цепочку ответов наподобие условных рефлексов, описанных Иваном Павловым: вид пищи вызывает у собаки мысли о еде, что, в свою очередь, вызывает слюноотделение. Но такая рефлекторная цепочка не требует никакого сознания. Она работает как линия розлива на заводе Coca-Cola: в бутылку наливается напиток, надевается крышка, приклеивается этикетка. Вряд ли кто-то будет утверждать, что у этой машины есть сознание.
Проблема еще больше усложняется тем, что значительная часть вычислений производится мозгом на бессознательном уровне. Во время езды на мотоцикле никто не говорит себе что-нибудь вроде: «На этом повороте мне нужно наклониться к внутреннему радиусу на столько-то градусов». Возможно, вы можете сформулировать это правило словами (а возможно, и нет), но вам не нужно каждый раз сознательно концентрировать на нем внимание, когда вы едете по извилистой дороге. Очень наглядно разницу между сознательным и бессознательным обучением демонстрирует случай знаменитого пациента Г.М. После удаления части мозга в целях ослабления эпилептических приступов этот человек утратил способность формировать новые сознательные воспоминания; например, он не запоминал прочитанное и мог читать один и тот же журнал снова и снова. Можно сказать, что его сознательная жизнь закончилась в день операции. Однако он по-прежнему был способен овладевать простыми моторными навыками, хотя и не мог ничего о них сказать и не помнил, как им научился.
Хебб не углублялся в проблему сознания. Он предполагал, что оно каким-то образом заключено в широко распределенной активности головного мозга, в одновременном возбуждении множества фазовых последовательностей. Кристоф Кох, много размышляющий о природе сознания, считает, что «сознание является фундаментальным, элементарным свойством живой материи», и признает неизбежно вытекающее из этого следствие, что «любая система взаимодействующих частей обладает некоторой мерой разумности». Это, помимо прочего, приводит нас к вопросу о сознании животных. Есть ли сознание у собаки? Многие считают, что да. А у червя C. Elegans с его 305 нейронами? Возможно, только совсем чуть-чуть. А как насчет дрозофилы со 135 000 нейронов или медузы с ее распределенной нейронной сетью? Согласно Хеббу, Коху и многим другим, сознание у млекопитающих возникает только тогда, когда их распределенные церебральные системы работают согласованным образом; соответственно сознание исчезает, когда более примитивные центры, контролирующие кору головного мозга, повреждаются или отключают функционирование коры (как во время сна). Другими словами, исследователи сходятся на том, что сознание зависит от неких крупных мозговых структур. Но какие именно нейронные цепи содержат сознание и какая именно нейрональная активность этих цепей порождает сознание? Можно сказать, что некая фазовая последовательность наивысшего порядка, распространившаяся по всей нейронной сети мозга через все ее внутренние границы, – это и есть сознание, своего рода эмерджентное свойство системы. Но такое «объяснение» ничего не объясняет; оно лишь переименовывает проблему. Возможно, сознание – это языковой конструкт, подобный парадоксам Зенона. Или же это новое свойство материи, аналогичное массе, как это постулирует Кох. К сожалению, наше внутреннее ощущение сознания не предлагает никаких зацепок, никакого наблюдательного места, с которого можно было бы изучить этот феномен. Сознание имманентно субъективно, оно существует только в конкретном человеке и нигде больше. Я боюсь, что сознание непознаваемо по своей природе. «Когда я вижу яблоко, – сказал однажды Дж. Э. Мур философу Бертрану Расселу, – как узнать, вижу ли я его таким же красным, как вы?» Насколько мне известно, никто еще не дал убедительного ответа на этот вопрос.
Глоссарий
HOG-изображение: карта, показывающая расположение градиентов яркости и их направление на изображении (гистограммы направленных градиентов).
Аксон: тонкий отросток, выходной элемент нейрона. В большинстве случаев сигналы идут от тела клетки к аксону и через синапс передаются на другой нейрон.
Альфа-клетка: структурный и функциональный тип ганглионарных клеток сетчатки с транзиторным on– или off-ответом на свет.
Амакриновая клетка: тип интернейронов сетчатки. Амакриновые клетки получают вход от биполярных клеток и других амакриновых клеток и передают выход на биполярные клетки, другие амакриновые клетки и ганглионарные клетки.
Ансамбль клеток: группа нейронов мозга, которые соединяются усиленными синаптическими связями в результате одновременной активации.
Ацетилхолин: органическое соединение, широко используемое в нервной системе в качестве нейромедиатора для связи между нейронами и между нейронами и мышечными клетками.
Бета-клетка: тип ганглионарных клеток сетчатки с устойчивым on– или off-ответом на свет и меньшими по размеру рецептивными полями, чем у альфа-клеток.
Биполярная клетка: тип интернейронов сетчатки. Эти кондукторные нейроны получают вход от фоторецепторных клеток в наружном слое сетчатки и передают выход на амакриновые и ганглионарные клетки во внутреннем слое.
Выделение краев: операция обработки изображений, которая увеличивает локальную разницу в интенсивности сигнала на границах визуальных объектов.
Ганглионарная клетка сетчатки: тип нейронов сетчатки, который получает входные сигналы от других нейронов (биполярных и амакриновых клеток) и передает выходные сигналы в мозг. Соединяясь вместе, аксоны ганглионарных клеток образуют зрительный нерв.
Гиппокамп: структура лимбической системы мозга, которая нравится экспериментаторам из-за стереотипности и доступности ее нейронов.
Горизонтальная клетка: интернейрон в наружном слое сетчатки; принимает входные сигналы от палочковых и колбочковых фоторецепторов и передает выходные сигналы обратно на те же рецепторы, а также на биполярные клетки.
Дендрит: тонкий разветвленный отросток, выходящий из клеточного тела нейрона. Традиционно считался входным элементом нейрона, но сейчас известно, что в некоторых случаях дендрит может быть и выходным элементом.
Долговременная потенциация (ДП): устойчивое усиление синаптической связи между двумя нейронами после всплеска активности.
Дофамин: химическое вещество, используемое в качестве нейромедиатора многими синапсами в головном мозге и сетчатке.
Закон Мура: эмпирическое наблюдение, согласно которому количество транзисторов в процессорах и соответственно вычислительная мощность компьютеров удваивается каждые 18 месяцев.
Избыточность: в теории информации под избыточностью понимается частичное совпадение данных в источнике информации. Когда мы говорим о зрении, избыточность означает, что некоторая точка на естественном видимом объекте с высокой вероятностью будет иметь те же характеристики, что и соседние точки.
Иммуноцитохимия: метод локализации белков в мозговой и других тканях; зависит от наличия антител к исследуемому белку.
Клетка с транзиторным ответом: в сенсорных системах нейрон, который реагирует на появление стимула, но затем снижает свою активность.
Клетка с устойчивым ответом: в сенсорных системах нейрон, который сохраняет возбуждение на протяжении всего времени действия стимула.
Кора: внешний слой серого вещества, покрывающий полушария головного мозга. Кора связана с подкорковыми структурами множеством реципрокных аксональных путей; задействована в сенсорном восприятии и высших психических функциях.
Латеральное коленчатое тело (ЛКТ): ядро, расположенное на дорсолатеральной поверхности таламуса; главная промежуточная станция на пути передачи зрительной информации от сетчатки в кору мозга.
Магнитно-резонансная томография (МРТ): способ получения на компьютере изображения внутренних органов и тканей, основанный на индуцировании ядерного магнитного резонанса в атомах (с помощью электромагнитных волн) и измерении их отклика.
Нейром: перечень всех типов клеток нервной структуры.
Нейромедиатор: химическое вещество, используемое для передачи возбуждения или торможения через синапсы.
Обратное распространение: метод настройки весовых коэффициентов соединений в нейронной сети, получающей инструкции от учителя.
Палочки и колбочки: два типа фоторецепторных клеток в сетчатке млекопитающих.
Постсинаптический нейрон: нейрон, получающий выходной сигнал от другого нейрона через синаптический контакт.
Потенциал действия: короткий электрический сигнал «все или ничего», генерируемый нейроном. Также известен как спайк. Потенциалы действия являются электрическим компонентом нервного импульса, передающегося от нейрона к нейрону.
Простая клетка: корковый нейрон, который лучше всего реагирует на линию или край определенной ориентации; в отличие от сложной клетки требует, чтобы изображение падало на строго определенную узкую область сетчатки.
Рецептивное поле: участок сенсорной поверхности, воздействие на который приводит к возбуждению или торможению конкретного нейрона. Изначально термин относился только к пространству, но теперь применяется и к другим признакам стимула, таким как движение, края и т. д., – например, можно сказать «чувствительное к направлению рецептивное поле».
Синапс Хебба: синапс, который ведет себя в соответствии с постулатом Дональда Хебба, то есть усиливается при одновременном возбуждении пре– и постсинаптического нейронов.
Синапс: место контакта двух нейронов, через которое могут передаваться возбуждающие или тормозящие сигналы. Можно сказать: «Биполярная клетка синаптирует с ганглионарной клеткой сетчатки».
Сложная клетка: тип нейронов зрительной коры. Сложные клетки лучше всего реагируют на линии и края определенной ориентации, которые могут располагаться в пределах довольно обширного рецептивного поля.
Спайк см. потенциал действия.
Средняя височная кора: зрительная область височной коры, специализирующаяся на обнаружении движущихся стимулов.
Флуоресцентный микроскоп: микроскоп, который получает изображение образца за счет излучаемого им света (флуоресценции). Образец облучается ярким светом с определенной длиной волны, что вызывает эмиссию света с другой длиной волны, который и улавливает микроскоп.
Фотозаполнение: совокупность методов с использованием диффундирующего маркера, который под воздействием света заполняет внутреннее клеточное пространство нейрона, включая аксон и дендриты, и делает его видимым среди неокрашенных соседних нейронов.
Фоторецепторные клетки: палочковые и колбочковые нейроны сетчатки, чувствительные к свету.
Циркадные часы: система мозга, отвечающая за поддержание циркадного ритма в телесных функциях, связанных с 24-часовым циклом сна и бодрствования.
Чувствительный к направлению нейрон: нейрон, сообщающий мозгу, в каком направлении движется стимул через его рецептивное поле.
Электронный микроскоп: микроскоп, который фокусирует на образце пучок электронов, а не фотонов (как световой микроскоп). Поскольку у электронов длина волны короче, чем у фотонов, разрешение электронной микроскопии выше, чем у световой.
Электроэнцефалограмма: запись электрической активности мозга с помощью электродов, расположенных вне мозга, обычно на поверхности скальпа.
Благодарности
Я хочу поблагодарить моих редакторов, в первую очередь уважаемых Бет Рапс и Лайзу Росс. Я познакомился с Лайзой, представляющей SDP Publishing, на курсах издательского дела в Гарварде, и она любезно направила меня к Бет. Бет впечатляющим образом улучшила мою книгу: каждый раз, когда я считал, что написал почти гениальную главу, она делала ее еще лучше. Если Бет сосредоточилась на форме, то Эрик Хенни из Basic Books сфокусировался на содержании. Он побуждал меня выйти за рамки сухой нейробиологии и приправить ее идеями из области восприятия, отчего эта книга стала куда интереснее.
Я многим обязан моим коллегам из Гарвардского университета и Массачусетского технологического института, щедро делившимся со мной своими идеями. Они знали, что я пишу книгу; многие прочитали отдельные главы и высказали свое мнение. Из благодарности к их открытости я не стал писать об их текущих исследованиях, результаты которых еще не были опубликованы. Я также благодарен всему сообществу исследователей сетчатки по всему миру, моим друзьям и критикам. За комментарии к отдельным главам я хочу поблагодарить Джудит Эймс, Марка Эймса, Ричарда Борна, Чинфей Чен, Филипа Крейвена, Дона Дондери, Марлу Феллер, Дэвида Гинти, Кристофера Харви, Габриэла Креймана, Маргарет Ливингстон, Стивена Мэсси, Питера Стерлинга, Энрику Стреттои, Уйгара Сюмбюла, Роя Вайза, Джереми Вулфа, которые избавили меня от многих ошибок.
Особая благодарность Элио Равиоле, моему другу и соавтору, который в свои 85 лет по-прежнему практически не покидает исследовательскую лабораторию. Спасибо Терренсу Сейновски за предварительный экземпляр его книги. Особая благодарность также Джеральду Ши за поддержку и полезный совет на ранней стадии. Эта книга не придерживается обычного научно-популярного формата, и я благодарен Джиму Левину из Levine Greenberg Rostan Literary Agency за то, что он дал мне шанс. Джим решил, что этот проект заслуживает внимания, и познакомил меня с вышеупомянутыми людьми в Basic Books.
И последнее по очередности, но не по важности: я хочу сказать спасибо моей жене Джин за комментарии к рукописям, за понимание, с которым она относилась к моей работе над книгой, и, самое главное, за то, что она всегда остается собой.
Библиография
ОБЩАЯ ОЗНАКОМИТЕЛЬНАЯ ЛИТЕРАТУРА
Ackerman, D. (1995). A natural history of the senses. New York: Vintage. [Акерман Д. Всеобщая история чувств. – М.: Колибри, 2018.] «Чувства доставляют в мозг информацию, рассыпанную на мельчайшие осколки, как пазл. Когда достаточное количество осколков складываются в нужном порядке, мозг говорит: «Корова. Я вижу корову». Изысканная книга о сенсорных системах, сочетающая точную науку и поэтический взгляд.
Dowling, J. E. (2012). The retina: An approachable part of the brain. Cambridge, MA: Harvard University Press. Классический учебник, на котором учатся уже несколько поколений студентов, подробно рассказывает о сетчатке и, в частности, о функциях интернейронов. Первое издание 1987 г.; обновленное 2012 г.
Hubel, D. (1988). Eye, brain, and vision. New York: W. H. Freeman. Обзор научной работы Хьюбела на уровне журнала Scientific American. Отточенная манера изложения, глубокая мысль за каждым словом. Не пытайтесь читать слишком быстро.
Masland, R. H. (2001). The fundamental plan of the retina. Nature Neuroscience, 4, 877–886. Попытка преодолеть видовые различия и выделить базовые организационные принципы, лежащие в основе работы и устройства сетчаток всех млекопитающих.
Rodieck, R. W. (1998). The first steps in seeing. Sunderland, MA: Sinauer. Боб Родик обладал поразительной широтой знаний в области зрения, от деталей фототрансдукции и анатомии ганглионарных клеток сетчатки до психофизики цветового зрения. Хотя книга немного устарела, она представляет собой кладезь фактов и глубоких размышлений о зрении. Прекрасно иллюстрирована самим Родиком с помощью его ассистента Тони Хоуна.
Wolfe, J. M., Kluender, K. R., Levi, D. M., Bartoshuk, L. M., Herz, R. S., Klatzky, R. L., & Merfeld, D. M. (2017). Sensation and perception, 5th ed. Sunderland, MA: Sinauer. Интересный и авторитетный учебник, охватывающий не только зрение, но и другие чувства.
ДОПОЛНИТЕЛЬНОЕ ЧТЕНИЕ
Много полезных материалов, особенно на тему машинного обучения, можно найти в интернете. Например, Дэвид Сильвер, руководитель команды, создавшей AlphaZero, в настоящее время публикует на YouTube замечательную серию лекций (свой курс в Университетском колледже Лондона). Но лет через пять они, скорее всего, будут удалены или заменены другими, поэтому указанные здесь ссылки окажутся устаревшими. Если вас заинтересовала эта тема, лучшего всего наберите в поисковике запрос «лекции по машинному обучению» и найдите наиболее актуальные.
Abraira, V. E., & Ginty, D. D. (2013). The sensory neurons of touch. Neuron, 79(4), 618–639.
Afraz, A., Boyden, E. S., & DiCarlo, J. J. (2015). Optogenetic and pharmacological suppression of spatial clusters of face neurons reveal their causal role in face gender discrimination. Proceedings of the National Academy of Sciences of the United States of America, 112(21), 6730–6735.
Albright, T. D. (1989). Centrifugal directional bias in the middle temporal visual area (MT) of the macaque. Visual Neuroscience, 2(2), 177–188.
Anzai, A., Peng, X., & Van Essen, D. C. (2007). Neurons in monkey visual area V2 encode combinations of orientations. Nature Neuroscience,10 (10), 1313–1321.
Arcaro, M. J., & Livingstone, M. S. (2017). A hierarchical, retinotopic proto-organization of the primate visual system at birth. Elife, 6.
Arcaro, M. J., Schade, P. F., Vincent, J. L., Ponce, C. R., & Livingstone, M. S. (2017). Seeing faces is necessary for face-domain formation. Nature Neuroscience, 20, 1404.
Arroyo, D. A., & Feller, M. B. (2016). Spatiotemporal features of retinal waves instruct the wiring of the visual circuitry. Frontiers in Neural Circuits, 10, 54.
Baden, T., Berens, P., Franke, K., Roman Roson, M., Bethge, M., & Euler, T. (2016). The functional diversity of retinal ganglion cells in the mouse. Nature, 529(7586), 345–350.
Ball, K., & Sekuler, R. (1982). A specific and enduring improvement in visual motion discrimination. Science, 218(4573), 697–698.
Ball, K., & Sekuler, R. (1987). Direction-specific improvement in motion discrimination. Vision Research, 27(6), 953–965.
Barone, I., Novelli, E., & Strettoi, E. (2014). Long-term preservation of cone photoreceptors and visual acuity in rd10 mutant mice exposed to continuous environmental enrichment. Molecular Vision, 20, 1545–1556.
Behrens, C., Schubert, T., Haverkamp, S., Euler, T., & Berens, P. (2016). Connectivity map of bipolar cells and photoreceptors in the mouse retina. Elife, 5.
Bell, A. J., & Sejnowski, T. J. (1997). The “independent components” of natural scenes are edge filters. Vision Research, 37(23), 3327–3338.
Bengio, Y. (2016). Machines who learn. Scientific American, 314(6), 46–51.
Bengio, Y., Courville, A., & Vincent, P. (2013). Representation learning: A review and new perspectives. IEEE Transactions on Pattern Analysis and Machine Intelligence, 35(8), 1798–1828.
Berry, K. P., & Nedivi, E. (2016). Experience-dependent structural plasticity in the visual system. Annual Review of Vision Science, 2, 17–35.
Besharse, J., & Bok, D. (Eds.). (2011). The retina and its disorders. San Diego, CA: Academic Press.
Blakemore, C., & Van Sluyters, R. C. (1975). Innate and environmental factors in the development of the kitten’s visual cortex. Journal of Physiology, 248(3), 663–716.
Bliss, T. V., & Lomo, T. (1973). Long-lasting potentiation of synaptic transmission in the dentate area of the anaesthetized rabbit following stimulation of the perforant path. Journal of Physiology, 232(2), 331–356.
Bojarski, M., Del Testa, D., Dworakowski, D., Firner, B., Flepp, B., Goyal, P., et al. (2016). End to end learning for self-driving cars. arXiv e-prints. Retrieved from https://ui.adsabs.harvard.edu/abs/2016arXiv160407316B.
Born, R. T., & Bradley, D. C. (2005). Structure and function of visual area MT. Annual Review of Neuroscience, 28, 157–189.
Boycott, B. B. (2001). Brian B. Boycott. In L. R. Squire (Ed.), The history of neuroscience in autobiography, volume 3. Cambridge, MA: Academic Press.
Boycott, B., & Wässle, H. (1999). Parallel processing in the mammalian retina: The Proctor Lecture. Investigative Ophthalmology and Visual Science, 40(7), 1313–1327.
Britten, K. H. (2008). Mechanisms of self-motion perception. Annual Review of Neuroscience, 31, 389–410.
Brown, R. E., & Milner, P. M. (2003). The legacy of Donald O. Hebb: More than the Hebb Synapse. Nature Reviews Neuroscience, 4, 1013.
Butts, D. A., Kanold, P. O., & Shatz, C. J. (2007). A burst-based “Hebbian” learning rule at retinogeniculate synapses links retinal waves to activity-dependent refinement. PLoS Biology, 5(3), e61.
Campbell, M. (2018). Mastering board games. Science, 362(6419), 1118.
Cang, J., Renteria, R. C., Kaneko, M., Liu, X., Copenhagen, D. R., & Stryker, M. P. (2005). Development of precise maps in visual cortex requires patterned spontaneous activity in the retina. Neuron, 48(5), 797–809.
Carandini, M. (2006). What simple and complex cells compute. Journal of Physiology, 577(Pt 2), 463–466.
Chang, L., & Tsao, D. Y. (2017). The code for facial identity in the primate brain. Cell, 169(6), 1013–1028 e1014.
Chapman, B., & Stryker, M. P. (1993). Development of orientation selectivity in ferret visual cortex and effects of deprivation. Journal of Neuroscience, 13(12), 5251–5262.
Chatterjee, R. (2015). Out of the darkness. Science, 350(6259), 372–375.
Chen, J., Yamahachi, H., & Gilbert, C. D. (2010). Experience-dependent gene expression in adult visual cortex. Cerebral Cortex, 20(3), 650–660.
Cohen, E., & Sterling, P. (1990). Demonstration of cell types among cone bipolar neurons of cat retina. Philosophical Transactions of the Royal Society of London. Series B, Biological Sciences, 330(1258), 305–321.
Coimbra, J. P., Marceliano, M. L., Andrade-da-Costa, B. L., & Yamada, E. S. (2006). The retina of tyrant flycatchers: Topographic organization of neuronal density and size in the ganglion cell layer of the great kiskadee Pitangus sulphuratus and the rusty margined flycatcher Myiozetetes cayanensis (Aves: Tyrannidae). Brain, Behavior and Evolution, 68(1), 15–25.
Costandi, M. (2009, February 10). Where are old memories stored in the brain? Scientific American. Retrieved from https://www.scientific american.com/article/the-memory-trace.
Crist, R. E., Kapadia, M. K., Westheimer, G., & Gilbert, C. D. (1997). Perceptual learning of spatial localization: Specificity for orientation, position, and context. Journal of Neurophysiology, 78(6), 2889–2894.
Dahne, S., Wilbert, N., & Wiskott, L. (2014). Slow feature analysis on retinal waves leads to V1 complex cells. PLOS Computational Biology, 10(5), e1003564.
Das, S. (2017). CNN architectures: LeNet, AlexNet, VGG, GoogLeNet, ResNet and more. Retrieved from https://medium.com/@sidereal/cnns-architectures-lenet-alexnet-vgg-googlenet-resnet-and-more-666091488df5.
Daw, N. (2006). Visual development (2nd ed.). New York: Springer.
Denk, W., Briggman, K. L., & Helmstaedter, M. (2012). Structural neurobiology: Missing link to a mechanistic understanding of neural computation. Nature Reviews Neuroscience, 13(5), 351–358.
DiCarlo, J. J., Zoccolan, D., & Rust, N. C. (2012). How does the brain solve visual object recognition? Neuron, 73(3), 415–434.
Dolan, T., & Fernandez-Juricic, E. (2010). Retinal ganglion cell topography of five species of ground-foraging birds. Brain, Behavior and Evolution, 75(2), 111–121.
Dormal, G., Lepore, F., & Collignon, O. (2012). Plasticity of the dorsal “spatial” stream in visually deprived individuals. Neural Plasticity, 2012, 659–687.
Dowling, J. E. (2012). The retina: An approachable part of the brain. Cambridge, MA: Harvard University Press.
Dowling, J. E., & Dowling, J. L. (2016). Vision: How it works and what can go wrong. Cambridge, MA: MIT Press.
Driscoll, L. N., Pettit, N. L., Minderer, M., Chettih, S. N., & Harvey, C. D. (2017). Dynamic reorganization of neuronal activity patterns in parietal cortex. Cell, 170(5), 986–999 e916.
Dvorak, D., Mark, R., & Reymond, L. (1983). Factors underlying falcon grating acuity. Nature, 303(5919), 729–730.
Eickhoff, S. B., Yeo, B. T. T., & Genon, S. (2018). Imaging-based parcellations of the human brain. Nature Reviews Neuroscience, 19(11), 672–686.
Eliot, V. (Ed.) (1971). The Waste Land: A Facsimile and Transcript of the Original Drafts. New York: Houghton Mifflin.
El-Shamayleh, Y., Kumbhani, R. D., Dhruv, N. T., & Movshon, J. A. (2013). Visual response properties of V1 neurons projecting to V2 in macaque. Journal of Neuroscience, 33(42), 16 594–16 605.
Escher, S. A., Tucker, A. M., Lundin, T. M., & Grabiner, M. D. (1998). Smokeless tobacco, reaction time, and strength in athletes. Medicine and Science in Sports and Exercise, 30(10), 1548–1551.
Espinosa, J. S., & Stryker, M. P. (2012). Development and plasticity of the primary visual cortex. Neuron, 75(2), 230–249.
Euler, T., Detwiler, P. B., & Denk, W. (2002). Directionally selective calcium signals in dendrites of starburst amacrine cells. Nature, 418(6900), 845–852.
Euler, T., & Wässle, H. (1995). Immunocytochemical identification of cone bipolar cells in the rat retina. Journal of Comparative Neurology, 361(3), 461–478.
Fisher, C., & Freiwald, W. A. (2015). Whole-agent selectivity within the macaque face-processing system. Proceedings of the National Academy of Sciences, 112(47), 14 717–14 722.
Fite, K. V., & Rosenfield-Wessels, S. (1975). A comparative study of deep avian foveas. Brain, Behavior and Evolution, 12(1–2), 97–115.
Fox, R., Lehmkuhle, S. W., & Westendorf, D. H. (1976). Falcon visual acuity. Science, 192(4236), 263–265.
Freeman, J., Field, G. D., Li, P. H., Greschner, M., Gunning, D. E., Mathieson, K., et al. (2015). Mapping nonlinear receptive field structure in primate retina at single cone resolution. Elife, 4.
Freeman, J., & Simoncelli, E. P. (2011). Metamers of the ventral stream. Nature Neuroscience, 14(9), 1195–1201.
Freiwald, W. A., & Tsao, D. Y. (2010). Functional compartmentalization and viewpoint generalization within the macaque face-processing system. Science, 330(6005), 845–851.
Gaffney, M. F., & Hodos, W. (2003). The visual acuity and refractive state of the American kestrel (Falco sparverius). Vision Research, 43(19), 2053–2059.
Gandhi, T. K., Ganesh, S., & Sinha, P. (2014). Improvement in spatial imagery following sight onset late in childhood. Psychological Science, 25(3), 693–701.
Gandhi, T. K., Singh, A. K., Swami, P., Ganesh, S., & Sinha, P. (2017). Emergence of categorical face perception after extended early-onset blindness. Proceedings of the National Academy of Sciences of the United States of America, 114(23), 6139–6143.
Gattass, R., Lima, B., Soares, J. G., & Ungerleider, L. G. (2015). Controversies about the visual areas located at the anterior border of area V2 in primates. Visual Neuroscience, 32, E019.
Gauthier, J. L., Field, G. D., Sher, A., Greschner, M., Shlens, J., Litke, A. M., & Chichilnisky, E. J. (2009). Receptive fields in primate retina are coordinated to sample visual space more uniformly. PLoS Biology, 7(4), e1000063.
Gauthier, I., & Tarr, M. J. (2016). Visual object recognition: Do we (finally) know more now than we did? Annual Review of Vision Science, 2, 377–396.
Gegenfurtner, K. R., Kiper, D. C., & Levitt, J. B. (1997). Functional properties of neurons in macaque area V3. Journal of Neurophysiology, 77(4), 1906–1923.
Geitgey, A. (2016, July 24). Machine learning is fun! Part 4: Modern face recognition with deep learning. Medium Artificial Intelligence. Retrieved from https://medium.com/@ageitgey/machine-learning-is-fun-part-4-modern-face-recognition-with-deep-learning-c3cffc121d78.
Ghim, M. M., & Hodos, W. (2006). Spatial contrast sensitivity of birds. Journal of Comparative Physiology. A, Neuroethology, Sensory, Neural, and Behavioral Physiology, 192(5), 523–534.
Ghose, G. M., Yang, T., & Maunsell, J. H. (2002). Physiological correlates of perceptual learning in monkey V1 and V2. Journal of Neurophysiology, 87(4), 1867–1888.
Gilbert, C. D., & Li, W. (2012). Adult visual cortical plasticity. Neuron, 75(2), 250–264.
Gollisch, T., & Meister, M. (2010). Eye smarter than scientists believed: Neural computations in circuits of the retina. Neuron, 65(2), 150–164.
Gopnik, A. (2017). Making AI more human. Scientific American, 316(6), 60–65.
Gopnik, A. (2019, February 22). Will A.I. ever be smarter than a four-year-old? Smithsonian.com. Retrieved from https://www.smithsonianmag.com/innovation/will-ai-ever-be-smarter-than-four-year-old-180971259.
Grady, C. L., Mondloch, C. J., Lewis, T. L., & Maurer, D. (2014). Early visual deprivation from congenital cataracts disrupts activity and functional connectivity in the face network. Neuropsychologia, 57, 122–139.
Gregory, R. L. (1997). Eye and brain: The psychology of seeing (5th ed.). Princeton, NJ: Princeton University Press.
Grens, K. (2014, November 1). A face to remember. The Scientist. Retrieved from https://www.the-scientist.com/cover-story/a-face-to-remember-36508.
Grimaldi, P., Saleem, K. S., & Tsao, D. (2016). Anatomical connections of the functionally defined “face patches” in the macaque monkey. Neuron, 90(6), 1325–1342.
Grimes, W. N., Songco-Aguas, A., & Rieke, F. (2018). Parallel processing of rod and cone signals: Retinal function and human perception. Annual Review of Vision Science, 4, 123–141.
Guillery, R. W. (2014). The lateral geniculate nucleus and pulvinar. In Werner, J. S., & Chalupa, L. M. (Eds.), The new visual neurosciences (pp. 257–283). Cambridge, MA: MIT Press.
Güntürkün, O. (1999). Sensory physiology: Vision. In G. Whittow (Ed.), Sturkie’s avian physiology (5th ed., pp. 1–19). Cambridge MA: Academic Press.
Hammond, P. (1974). Cat retinal ganglion cells: Size and shape of receptive field centres. Journal of Physiology, 242(1), 99–118.
Hebb, D. O. (1949). The organization of behavior: A neuropsychological theory. New York: Wiley.
Hebb, D. O. (1980). D. O. Hebb. In G. Lindzey (Ed.), A history of psychology in autobiography, vol. VII (pp. 273–303). San Francisco: W. H. Freeman.
Helmstaedter, M., Briggman, K. L., Turaga, S. C., Jain, V., Seung, H. S., & Denk, W. (2013). Connectomic reconstruction of the inner plexiform layer in the mouse retina. Nature, 500(7461), 168–174.
Hinton, G., Deng, L., Yu, D., Dahl, G. E., Mohamed, A., Jaitly, N., et al. (2012). Deep neural networks for acoustic modeling in speech recognition: The shared views of four research groups. IEEE Signal Processing Magazine, 29(6), 82–97.
Hodos, W., Ghim, M. M., Potocki, A., Fields, J. N., & Storm, T. (2002). Contrast sensitivity in pigeons: A comparison of behavioral and pattern ERG methods. Documenta Ophthalmologica, 104(1), 107–118.
Holcombe, A. O. (2010). Binding problem. In E. B. Goldstein (Ed.), Encyclopedia of Perception (pp. 206–208). Thousand Oaks, CA: SAGE Publications.
Hubel, D. (1995). Eye, brain, and vision. New York: Scientific American.
Huberman, A. D., Feller, M. B., & Chapman, B. (2008). Mechanisms underlying development of visual maps and receptive fields. Annual Review of Neuroscience, 31, 479–509.
Hung, C. P., Kreiman, G., Poggio, T., & DiCarlo, J. J. (2005). Fast readout of object identity from macaque inferior temporal cortex. Science, 310(5749), 863–866.
Ings, S. (2007). The eye: A natural history. London: Bloomsbury.
Inzunza, O., Bravo, H., Smith, R. L., & Angel, M. (1991). Topography and morphology of retinal ganglion cells in Falconiforms: A study on predatory and carrion-eating birds. Anatomical Record, 229 (2), 271–277.
Issa, E. B., & DiCarlo, J. J. (2012). Precedence of the eye region in neural processing of faces. Journal of Neuroscience, 32(47), 16666–16682.
Jacoby, J., & Schwartz, G. W. (2017). Three small-receptive-field ganglion cells in the mouse retina are distinctly tuned to size, speed, and object motion. Journal of Neuroscience, 37(3), 610–625.
Kalloniatis, M., & Luu, C. (2007). Visual acuity. Webvision. Retrieved from https://webvision.med.utah.edu/book/part-viii-psychophysics-of-vision/visual-acuity.
Kandel, E. (2001). Nobel Lecture: The molecular biology of memory storage: A dialog between genes and synapses. Bioscience Reports, 21, 565–611.
Kaneko, M., & Stryker, M. P. (2017). Homeostatic plasticity mechanisms in mouse V1. Philosophical Transactions of the Royal Society of London. Series B, Biological Sciences, 372(1715).
Kiani, R., Esteky, H., Mirpour, K., & Tanaka, K. (2007). Object category structure in response patterns of neuronal population in monkey inferior temporal cortex. Journal of Neurophysiology, 97(6), 4296–4309.
Kienker, P. K., Sejnowski, T. J., Hinton, G. E., & Schumacher, L. E. (1986). Separating figure from ground with a parallel network. Perception, 15(2), 197–216.
Kirkby, L. A., Sack, G. S., Firl, A., & Feller, M. B. (2013). A role for correlated spontaneous activity in the assembly of neural circuits. Neuron, 80(5), 1129–1144.
Koch, C. (2012). Consciousness: Confessions of a romantic reductionist. Cambridge, MA: MIT Press.
Kolb, H. (2006). Facts and figures concerning the human retina. Webvision. Retrieved from https://webvision.med.utah.edu/book/part-xiii-facts-and-figures-concerning-the-human-retina.
Kornblith, S., & Tsao, D. Y. (2017). How thoughts arise from sights: Inferotemporal and prefrontal contributions to vision. Current Opinion in Neurobiology, 46, 208–218.
Krauzlis, R. J., Lovejoy, L. P., & Zenon, A. (2013). Superior colliculus and visual spatial attention. Annual Review of Neuroscience, 36, 165–182.
Krieger, B., Qiao, M., Rousso, D. L., Sanes, J. R., & Meister, M. (2017). Four alpha ganglion cell types in mouse retina: Function, structure, and molecular signatures. PLoS One, 12(7), e0180091.
Kumano, H., & Uka, T. (2013). Neuronal mechanisms of visual perceptual learning. Behavioural Brain Research, 249, 75–80.
Lashley, K. S. (1950). In search of the engram. Physiological mechanisms in animal behavior (Society’s Symposium IV) (pp. 454–482). Oxford, UK: Academic Press.
LeCun, Y., Bengio, Y., & Hinton, G. (2015). Deep learning. Nature, 521(7553), 436–444.
Lewis-Kraus, G. (2016, December 14). The great A.I. awakening. New York Times Magazine. Retrieved from https://www.nytimes.com/2016/12/14/magazine/the-great-ai-awakening.html.
Li, S. Z., & Jain, A. (Eds.). (2011). Handbook of face recognition (2nd ed.). New York: Springer.
Lindsey, J., Ocko, S. A., Ganguli, S., & Deny, S. (2019). A unified theory of early visual representations from retina to cortex through anatomically constrained deep CNNs. arXiv e-prints. Retrieved from https://ui.adsabs.harvard.edu/abs/2019arXiv190100945L.
Litvina, E. Y., & Chen, C. (2017). Functional convergence at the retinogeniculate synapse. Neuron, 96(2), 330–338 e335.
Liu, L., She, L., Chen, M., Liu, T., Lu, H. D., Dan, Y., & Poo, M. M. (2016). Spatial structure of neuronal receptive field in awake monkey secondary visual cortex (V2). Proceedings of the National Academy of Sciences of the United States of America, 113(7), 1913–1918.
Liu, Y. S., Stevens, C. F., & Sharpee, T. O. (2009). Predictable irregularities in retinal receptive fields. Proceedings of the National Academy of Sciences of the United States of America, 106(38), 16 499–16 504.
Livingstone, M. S., Pack, C. C., & Born, R. T. (2001). Two-dimensional substructure of MT receptive fields. Neuron, 30(3), 781–793.
Livingstone, M. S., Vincent, J. L., Arcaro, M. J., Srihasam, K., Schade, P. F., & Savage, T. (2017). Development of the macaque face-patch system. Nature Communications, 8, 14 897.
MacNeil, M. A., Heussy, J. K., Dacheux, R. F., Raviola, E., & Masland, R. H. (1999). The shapes and numbers of amacrine cells: Matching of photofilled with Golgi-stained cells in the rabbit retina and comparison with other mammalian species. Journal of Comparative Neurology, 413, 305–326.
MacNeil, M. A., Heussy, J. K., Dacheux, R. F., Raviola, E., & Masland, R. H. (2004). The population of bipolar cells in the rabbit retina. Journal of Comparative Neurology, 472, 73–86.
Margolis, D. J., Lutcke, H., Schulz, K., Haiss, F., Weber, B., Kugler, S., et al. (2012). Reorganization of cortical population activity imaged throughout long-term sensory deprivation. Nature Neuroscience, 15(11), 1539–1546.
Martersteck, E. M., Hirokawa, K. E., Evarts, M., Bernard, A., Duan, X., Li, Y., et al. (2017). Diverse central projection patterns of retinal ganglion cells. Cell Reports, 18(8), 2058–2072.
Martin, P., & Masland, R. H. (2007). Essay: The unsolved mystery of vision. Current Biology 18: R577–R583.
Masland, R. H. (2001). Neuronal diversity in the retina. Current Opinion in Neurobiology, 11, 431–436.
Masland, R. H. (2012). The neuronal organization of the retina. Neuron, 76, 266–280.
McGuire, B. A., Stevens, J. K., & Sterling, P. (1984). Microcircuitry of bipolar cells in cat retina. Journal of Neuroscience, 4(12), 2920–2938.
McKyton, A., Ben-Zion, I., Doron, R., & Zohary, E. (2015). The limits of shape recognition following late emergence from blindness. Current Biology, 25(18), 2373–2378.
McMahan, U. (1990). Steve: Remembrances of Stephen W. Kuffler. Sunderland, MA: Sinauer Associates.
McManus, J. N., Li, W., & Gilbert, C. D. (2011). Adaptive shape processing in primary visual cortex. Proceedings of the National Academy of Sciences of the United States of America, 108(24), 9739–9746.
Meyers, E. M., Borzello, M., Freiwald, W. A., & Tsao, D. (2015). Intelligent information loss: The coding of facial identity, head pose, and non-face information in the macaque face patch system. Journal of Neuroscience, 35(18), 7069–7081.
Moeller, S., Crapse, T., Chang, L., & Tsao, D. Y. (2017). The effect of face patch microstimulation on perception of faces and objects. Nature Neuroscience, 20(5), 743–752.
Montaсez, A. (2016, May 20). Unveiling the hidden layers of deep learning. SA Visual, a blog in Scientific American. Retrieved from https://blogs.scientificamerican.com/sa-visual/unveiling-the-hidden-layers-of-deep-learning.
Moore, B. A., Tyrrell, L. P., Pita, D., Bininda-Emonds, O. R. P., & Fernбndez– Juricic, E. (2017). Does retinal configuration make the head and eyes of foveate birds move? Scientific Reports, 7, 38 406.
Moore, B. D., Kiley, C. W., Sun, C., & Usrey, W. M. (2011). Rapid plasticity of visual responses in the adult lateral geniculate nucleus. Neuron, 71(5), 812–819.
Morgan, J. L., Berger, D. R., Wetzel, A. W., & Lichtman, J. W. (2016). The fuzzy logic of network connectivity in mouse visual thalamus. Cell, 165(1), 192–206.
Movshon, J. A., Lisberger, S. G., & Krauzlis, R. J. (1990). Visual cortical signals supporting smooth pursuit eye movements. Cold Spring Harbor Symposia on Quantitative Biology, 55, 707–716.
Movshon, J. A., & Newsome, W. T. (1996). Visual response properties of striate cortical neurons projecting to area MT in macaque monkeys. Journal of Neuroscience, 16(23), 7733–7741.
Movshon, J. A., & Simoncelli, E. P. (2014). Representation of naturalistic image structure in the primate visual cortex. Cold Spring Harbor Symposia on Quantitative Biology, 79, 115–122.
Ohki, K., Chung, S., Ch’ng, Y. H., Kara, P., & Reid, R. C. (2005). Functional imaging with cellular resolution reveals precise microarchitecture in visual cortex. Nature, 433(7026), 597–603.
O’Keefe, J. (2014, December 7). Spatial cells in the hippocampal formation. Nobel Lecture.
O’Keefe, J., & Dostrovsky, J. (1971). The hippocampus as a spatial map: Preliminary evidence from unit activity in the freely-moving rat. Brain Research, 34(1), 171–175.
Olshausen, B. A., & Field, D. J. (1996). Natural image statistics and efficient coding. Network, 7(2), 333–339.
O’Rourke, C. T., Hall, M. I., Pitlik, T., & Fernandez-Juricic, E. (2010). Hawk eyes I: Diurnal raptors differ in visual fields and degree of eye movement. PLoS One, 5(9), e12802.
Pack, C. C., & Born, R. T. (2004). Responses of MT neurons to barber pole stimuli. Journal of Vision, 4(859).
Pack, C. C., & Born, R. (2010). Cortical mechanisms for the integration of visual motion. In R. H. Masland, T. D. Albright, G. M. Shephard, & E. P. Gardner (Eds.), The senses: A comprehensive reference, vol. 2 (pp. 189–218). San Diego, CA: Academic Press.
Pack, C. C., Gartland, A. J., & Born, R. T. (2004). Integration of contour and terminator signals in visual area MT of alert macaque. Journal of Neuroscience, 24(13), 3268–3280.
Peron, S. P., Freeman, J., Iyer, V., Guo, C., & Svoboda, K. (2015). A cellular resolution map of barrel cortex activity during tactile behavior. Neuron, 86(3), 783–799.
Phillips, P. J., Grother, P., Michaels, R. J., Balackburn, D. M., Tabassi, E., & Bone, M. (2003). Face recognition vendor test 2002: Evaluation report (6965). Retrieved from https://nvlpubs.nist.gov/nistpubs/Legacy/IR/nistir6965.pdf.
Ponce, C. R., Hartmann, T. S., & Livingstone, M. S. (2017). Endstopping predicts curvature tuning along the ventral stream. Journal of Neuroscience, 37(3), 648–659.
Pritchard, R. M., Heron, W., & Hebb, D. O. (1960). Visual perception approached by the method of stabilized images. Canadian Journal of Experimental Psychology, 14, 67–77.
Protti, D. A., Flores-Herr, N., Li, W., Massey, S. C., & Wässle, H. (2005). Light signaling in scotopic conditions in the rabbit, mouse and rat retina: A physiological and anatomical study. Journal of Neurophysiology, 93(6), 3479–3488.
Quiroga, R. Q., Reddy, L., Kreiman, G., Koch, C., & Fried, I. (2005). Invariant visual representation by single neurons in the human brain. Nature, 435(7045), 1102–1107.
Raiguel, S., Van Hulle, M. M., Xiao, D. K., Marcar, V. L., & Orban, G. A. (1995). Shape and spatial distribution of receptive fields and antagonistic motion surrounds in the middle temporal area (V5) of the macaque. European Journal of Neuroscience, 7(10), 2064–2082.
Reid, R. C., & Alonso, J. M. (1995). Specificity of monosynaptic connections from thalamus to visual cortex. Nature, 378(6554), 281–284.
Reid, R. C., & Usrey, W. M. (2013). Vision. In L. Squire, D. Berg, F. E. Bloom, S. du Lac, A. Ghosh, & N. C. Spitzer (Eds.), Fundamental neuroscience (4th ed., pp. 577–595). Oxford, UK: Academic Press.
Reymond, L. (1985). Spatial visual acuity of the eagle Aquila audax: A behavioural, optical and anatomical investigation. Vision Research, 25(10), 1477–1491.
Reymond, L. (1987). Spatial visual acuity of the falcon, Falco berigora: A behavioural, optical and anatomical investigation. Vision Research, 27(10), 1859–1874.
Richert, M., Albright, T. D., & Krekelberg, B. (2013). The complex structure of receptive fields in the middle temporal area. Frontiers in Systems Neuroscience, 7, 2.
Riesenhuber, M., & Poggio, T. (1999). Hierarchical models of object recognition in cortex. Nature Neuroscience, 2(11), 1019–1025.
Roe, A. W., Chelazzi, L., Connor, C. E., Conway, B. R., Fujita, I., Gallant, J. L., et al. (2012). Toward a unified theory of visual area V4. Neuron, 74(1), 12–29.
Roelfsema, P. R., & Holtmaat, A. (2018). Control of synaptic plasticity in deep cortical networks. Nature Reviews Neuroscience, 19(3), 166–180.
Roman Roson, M., Bauer, Y., Kotkat, A. H., Berens, P., Euler, T., & Busse, L. (2019). Mouse dLGN receives functional input from a diverse population of retinal ganglion cells with limited convergence. Neuron, 102(2), 462–476 e468.
Rompani, S. B., Mullner, F. E., Wanner, A., Zhang, C., Roth, C. N., Yonehara, K., & Roska, B. (2017). Different modes of visual integration in the lateral geniculate nucleus revealed by single-cell-initiated transsynaptic tracing. Neuron, 93(4), 767–777.
Rose, T., & Bonhoeffer, T. (2018). Experience-dependent plasticity in the lateral geniculate nucleus. Current Opinion in Neurobiology, 53, 22–28.
Roska, B. (2019). The first steps in vision: Cell types, circuits, and repair. EMBO Molecular Medicine, 11(3).
Rossion, B., & Taubert, J. (2017). Commentary: The code for facial identity in the primate brain. Frontiers in Human Neuroscience, 11, 550.
Ruggeri, M., Major, J. C. Jr., McKeown, C., Knighton, R. W., Puliafito, C. A., & Jiao, S. (2010). Retinal structure of birds of prey revealed by ultra-high resolution spectral-domain optical coherence tomography. Investigative Ophthalmology and Visual Science, 51(11), 5789–5795.
Sagi, D. (2011). Perceptual learning in vision research. Vision Research, 51(13), 1552–1566.
Sanes, J. R., & Masland, R. H. (2015). The types of retinal ganglion cells: Current status and implications for neuronal classification. Annual Review of Neuroscience, 38, 221–246.
Scholl, B., & Priebe, N. J. (2015). Neuroscience: The cortical connection. Nature, 518(7539), 306–307.
Seabrook, T. A., Burbridge, T. J., Crair, M. C., & Huberman, A. D. (2017). Architecture, function, and assembly of the mouse visual system. Annual Review of Neuroscience, 40, 499–538.
Sejnowski, T. J. (2018). The deep learning revolution: Artificial intelligence meets human intelligence. Cambridge, MA: MIT Press.
Seung, S. (2012). Connectome: How the brain’s wiring makes us who we are. New York: Houghton Mifflin Harcourt.
Shadlen, M. N., & Movshon, J. A. (1999). Synchrony unbound: A critical evaluation of the temporal binding hypothesis. Neuron, 24(1), 67–77, 111–125.
Sharma, J., Angelucci, A., & Sur, M. (2000). Induction of visual orientation modules in auditory cortex. Nature, 404(6780), 841–847.
Sheikh, K. (2017, June 1). How we save face – Researchers crack the brain’s facial-recognition code. Scientific American. Retrieved from https://www.scientificamerican.com/article/how-we-save-face-researchers-crack-the-brains-facial-recognition-code.
Shekhar, K., Lapan, S. W., Whitney, I. E., Tran, N. M., Macosko, E. Z., Kowalczyk, M., et al. (2016). Comprehensive classification of retinal bipolar neurons by single-cell transcriptomics. Cell, 166(5), 1308–1323 e1330.
Sherman, S. M., & Guillery, R. W. (2013). Functional connections of cortical areas. Cambridge, MA: MIT Press.
Silver, D., Hubert, T., Schrittwieser, J., Antonoglou, I., Lai, M., Guez, A., et al. (2018). A general reinforcement learning algorithm that masters chess, shogi, and Go through self-play. Science, 362 (6419), 1140–1144.
Sincich, L. C., Horton, J. C., & Sharpee, T. O. (2009). Preserving information in neural transmission. Journal of Neuroscience, 29(19), 6207–6216.
Sinha, P. (2013). Once blind and now they see. Scientific American, 309(1), 48–55.
Smolyanskaya, A., Haefner, R. M., Lomber, S. G., & Born, R. T. (2015). A modality-specific feedforward component of choice-related activity in MT. Neuron, 87(1), 208–219.
Solomon, S. G., Tailby, C., Cheong, S. K., & Camp, A. J. (2010). Linear and nonlinear contributions to the visual sensitivity of neurons in primate lateral geniculate nucleus. Journal of Neurophysiology, 104(4), 1884–1898.
Srihasam, K., Vincent, J. L., & Livingstone, M. S. (2014). Novel domain formation reveals proto-architecture in inferotemporal cortex. Nature Neuroscience, 17(12), 1776–1783.
Stevens, C. F. (1998). Neuronal diversity: Too many cell types for comfort? Current Biology, 8(20), R708–710.
Stokkan, K. A., Folkow, L., Dukes, J., Neveu, M., Hogg, C., Siefken, S., et al. (2013). Shifting mirrors: Adaptive changes in retinal reflections to winter darkness in Arctic reindeer. Proceedings of the Royal Society B: Biological Sciences, 280(1773), 2013–2451.
Strogatz, S. (2018, December 26). One giant step for a chess-playing machine. New York Times. Retrieved from https://www.nytimes.com/2018/12/26/science/chess-artificial-intelligence.html.
Strom, R. C. (1999). Genetic and environmental control of retinal ganglion cell variation. Chapter 4 in Genetic analysis of variation in neuron number, PhD diss., University of Tennessee Health Science Center, Memphis, Tennessee. Retrieved from www.nervenet.org/papers/strom99/Chapter4.html.
Sumbul, U., Song, S., McCulloch, K., Becker, M., Lin, B., Sanes, J. R., et al. (2014). A genetic and computational approach to structurally classify neuronal types. Nature Communications, 5, 3512.
Suresh, V., Ciftcioglu, U. M., Wang, X., Lala, B. M., Ding, K. R., Smith, W. A., et al. (2016). Synaptic contributions to receptive field structure and response properties in the rodent lateral geniculate nucleus of the thalamus. Journal of Neuroscience, 36(43), 10 949–10 963.
Tanaka, K. (1983). Cross-correlation analysis of geniculostriate neuronal relationships in cats. Journal of Neurophysiology, 49(6), 1303–1318.
Tanaka, K. (1985). Organization of geniculate inputs to visual cortical cells in the cat. Vision Research, 25(3), 357–364.
Tang, S., Lee, T. S., Li, M., Zhang, Y., Xu, Y., Liu, F., et al. (2018). Complex pattern selectivity in macaque primary visual cortex revealed by large-scale two-photon imaging. Current Biology, 28(1), 38–48 e33.
Thompson, A., Gribizis, A., Chen, C., & Crair, M. C. (2017). Activity-dependent development of visual receptive fields. Current Opinion in Neurobiology, 42, 136–143.
Tien, N. W., Pearson, J. T., Heller, C. R., Demas, J., & Kerschensteiner, D. (2015). Genetically identified suppressed-by-contrast retinal ganglion cells reliably signal self-generated visual stimuli. Journal of Neuroscience, 35(30), 10 815–10 820.
Tonegawa, S., Liu, X., Ramirez, S., & Redondo, R. (2015). Memory engram cells have come of age. Neuron, 87(5), 918–931.
Tootell, R. B., Reppas, J. B., Dale, A. M., Look, R. B., Sereno, M. I., Malach, R., et al. (1995). Visual motion aftereffect in human cortical area MT revealed by functional magnetic resonance imaging. Nature, 375(6527), 139–141.
Tsao, D. (2014). The macaque face patch system: A window into object representation. Cold Spring Harbor Symposia on Quantitative Biology, 79, 109–114.
Tsao, D. Y., & Livingstone, M. S. (2008). Mechanisms of face perception. Annual Review of Neuroscience, 31, 411–437.
Tsodyks, M., & Gilbert, C. (2004). Neural networks and perceptual learning. Nature, 431(7010), 775–781.
Turner, M. H., Sanchez Giraldo, L. G., Schwartz, O., & Rieke, F. (2019). Stimulus– and goal-oriented frameworks for understanding natural vision. Nature Neuroscience, 22(1), 15–24.
Wagner, I. C. (2016). The integration of distributed memory traces. Journal of Neuroscience, 36(42), 10723–10725.
Wandell, B. A., & Smirnakis, S. M. (2009). Plasticity and stability of visual field maps in adult primary visual cortex. Nature Reviews Neuroscience, 10(12), 873–884.
Wang, H. X., & Movshon, J. A. (2016). Properties of pattern and component direction-selective cells in area MT of the macaque. Journal of Neurophysiology, 115(6), 2705–2720.
Wässle, H. (2002). Brian Blundell Boycott, 10 December 1924–22 April 2000. Biographical Memoirs of Fellows of the Royal Society, 48, 53–68.
Wässle, H., Grunert, U., Rohrenbeck, J., & Boycott, B. B. (1989). Cortical magnification factor and the ganglion cell density of the primate retina. Nature, 341(6243), 643–646.
Wässle, H., Puller, C., Muller, F., & Haverkamp, S. (2009). Cone contacts, mosaics, and territories of bipolar cells in the mouse retina. Journal of Neuroscience, 29(1), 106–117.
Watanabe, T., Nбсez, J. E., & Sasaki, Y. (2001). Perceptual learning without perception. Nature, 413(6858), 844–848.
Wathey, J. C., & Pettigrew, J. D. (1989). Quantitative analysis of the retinal ganglion cell layer and optic nerve of the barn owl Tyto alba. Brain, Behavior and Evolution, 33(5), 279–292.
Werner, J. S., & Chalupa, L. M. (Eds.). (2014). The new visual neurosciences. Cambridge, MA: The MIT Press.
Wiesel, T. N. (1982). Postnatal development of the visual cortex and the influence of environment. Nature, 299(5884), 583–591.
Wong, R. O., Meister, M., & Shatz, C. J. (1993). Transient period of correlated bursting activity during development of the mammalian retina. Neuron, 11(5), 923–938.
Wu, K. J. (2018, December 10). Google’s new A.I. is a master of games, but how does it compare to the human mind? Smithsonian.com. Retrieved from www.smithsonianmag.com/innovation/google-ai-deepminds-alphazero-games-chess-and-go-180970981.
Yamins, D. L., & DiCarlo, J. J. (2016). Using goal-driven deep learning models to understand sensory cortex. Nature Neuroscience, 19(3), 356–365.
Yamins, D. L. K., Hong, H., Cadieu, C. F., Solomon, E. A., Seibert, D., & DiCarlo, J. J. (2014). Performance-optimized hierarchical models predict neural responses in higher visual cortex. Proceedings of the National Academy of Sciences, 111(23), 8619–8624.
Zeng, H., & Sanes, J. R. (2017). Neuronal cell-type classification: Challenges, opportunities and the path forward. Nature Reviews Neuroscience, 18(9), 530–546.
Zhang, X., Zhao, J., & LeCun, Y. (2015). Character-level convolutional networks for text classification. In Cortes, C., Lawrence, N. D., Lee, D. D., Sugiyama, M., & Garnett, R. (Eds.), Advances in neural information processing systems 28. Red Hook, NY: Curran.
Zimmerman, A., Bai, L., & Ginty, D. D. (2014). The gentle touch receptors of mammalian skin. Science, 346(6212), 950–954.
Zuccolo, R. (2017, April 3). Self-driving cars – Advanced computer vision with opencv, finding lane lines. Retrieved from https://chatbotslife.com/self-driving-cars-advanced-computer-vision-with-opencv-finding-lane-lines-488a411b2c3d.
Источники иллюстраций
Иллюстрации, не указанные ниже, принадлежат автору и Хаобин Вану.
Глава 1. Три лица: фрагмент рекламного плаката New York City Ballet. Предоставлено Полом Кольником.
Глава 4. Биполярные клетки: рисунок Элио Равиолы.
Глава 5. Баскетболист: с сайта Pixabay@pexel.com; автор Кит Джонсон. Нейроны сетчатки: рисунок Ричарда Маслэнда и Ребекки Рокхилл.
Глава 6. Простая клетка: Hubel, D. (1988), Eye, brain, and vision, New York: Scientific American Library. Сложная клетка: Eye, brain, and vision, New York: Scientific American Library.
Глава 7. Мозг макаки: Yang, R. Visual areas of macaque monkey; http://fourier.eng.hmc.edu/e180/lectures/visualcortex/node6.html. Участки распознавания лиц в мозге мартышки: Hung, C. C., Yen, C. C., Ciuchta, J. L., Papoti, D., Bock, N. A., Leopold, D. A., & Silva, A. C. (2015), “Functional mapping of face-selective regions in the extrastriate visual cortex of the marmoset”, Journal of Neuroscience, 21(35), 1160–1172. Схематические лица: Tsao, D. (2014), “Detail from: The macaque face patch system: a window into object representation”, Cold Spring Harbor Symposia on Quantitative Biology, 79, 109–114.
Глава 10. HOG-изображение: https://medium.com/@ageitgey/machine-learning-is-fun-part-4-modern-face-recognition-with-deep-learning-c3cffc121d78. Схема говорящей нейросети: Rosenberg, C. R., and Sejnowski, T. (1987), “Parallel networks that learn to pronounce English text”, Journal of Complex Systems, 1, 145–168. Классическая нейронная сеть: Bengio, Y. (2016), “Machines who learn”, Scientific American, 314, 46–51. Оригинальный рисунок Джена Кристиансена.
Глава 11. Связи в коре мозга: Felleman, D., and Van Essen, D. (1991), “Distributed hierarchical processing in the primate cerebral cortex”, Cerebral Cortex, 1, 1–47.
Глава 14. Проблема связывания: Sinha, P. (2014), “Once blind and now they see”, Scientific American, 309, 49–55. Изображение любезно предоставлено проектом Пракаша.
Об авторе

Ричард Маслэнд (1942–2019) был заслуженным профессором офтальмологии кафедры имени Дэвида Гленденинга Когана и профессором нейронаук в Гарвардской медицинской школе. Занимал пост вице-председателя по офтальмологическим исследованиям в Массачусетской клинике болезней глаз, уха, горла и носа при Гарвардском университете, являющейся крупнейшим в мире центром изучения зрения. Более 20 лет преподавал в Гарвардской медицинской школе курс нейробиологии, который дважды признавался лучшей учебной программой. Был членом Американской ассоциации содействия развитию науки (AAAS), исследователем в Медицинском институте Говарда Хьюза, лауреатом премии Проктора за научные достижения, премии Alcon Research и многих других. Работы Маслэнда открыли новые горизонты в изучении нейронных сетей сетчатки глаза.
Сноски
1
Каждый зрительный нерв состоит примерно из миллиона нервных волокон. Слуховой нерв – всего из 30 000. Эти два вида восприятия имеют разную основу. Зрение прежде всего основано на регистрации пространства – через распределение входящих сигналов по сетчатке. Слух регистрирует время – последовательность, в которой звуковые волны достигают ушей. Интересно, что временно́е разрешение слуховой системы намного лучше, чем у зрительной. Но проблема сегментации одинакова в обоих случаях: зрению нужно отделить объекты от окружающего фона; слуху – вычленить значимый звук из сопровождающего его шума.
(обратно)2
Одни нейромедиаторы возбуждают постсинаптический нейрон, другие тормозят; одни действуют очень быстро (за миллисекунды), другие довольно медленно (за секунды и даже десятки секунд). Во многих, но не во всех случаях нейрон может синтезировать только один вид нейромедиатора. Учитывая, что разные типы нейронов имеют тысячи различных паттернов связей с другими нейронами, дополнительное многообразие нейромедиаторов значительно увеличивает разнообразие вычислений, которые может выполнять мозг.
(обратно)3
На самом деле на поверхности головного мозга имеется полная карта поверхности тела, то есть каждой части тела соответствует свой участок на поверхности коры. В результате возбуждение нейронов в зоне большого пальца говорит мозгу, что что-то происходит с большим пальцем.
(обратно)4
Например, паттерн спайков, соответствующий букве А в азбуке Морзе, может сообщать мозгу, что сигнал поступает от быстро адаптирующегося тактильного афферентного нейрона.
(обратно)5
Дэвид Гинти. См.: Zimmerman, A., Bai, L., and Ginty, D. D. (2014), “The gentle touch receptors of mammalian skin”, Science, 346, 950–954; Abraira, V. E., and Ginty, D. D. (2013), “The sensory neurons of touch”, Neuron, 79(4), 618–639.
(обратно)6
Рассказ о Стивене Куффлере основан на личных воспоминаниях автора, а также на эссе из сборника: McMahan, U. J. (1990), Steve: Remembrances of Stephen W. Kuffler, Sunderland, MA: Sinauer.
(обратно)7
Переоборудованный и ярко раскрашенный школьный автобус, на котором члены коммуны хиппи «Веселые проказники» во главе с идеологом Кеном Кизи в 1960-х гг. путешествовали по США. – Прим. пер.
(обратно)8
Сделать это не так-то просто, что объясняет, почему авторы известной статьи пришли к ошибочному выводу, будто у ястребов поразительно острое зрение. Причина ошибки крылась в непродуманной поведенческой парадигме, которая использовалась для тестирования ястребиного зрения. См.: Gaffney, M. F., & Hodos, W. (2003), “The visual acuity and refractive state of the American kestrel (Falco sparverius)”, Vision Research, 43, 2053–2059.
(обратно)9
В действительности пустельга – птица семейства соколиных. – Прим. пер.
(обратно)10
Этот механизм и его открытие хорошо описаны в книге: Dowling, J. E. (2012), The retina: An approachable part of the brain, Cambridge, MA: Harvard University Press.
(обратно)11
Gollisch, T., & Meister, M. (2010), “Eye smarter than scientists believed: neural computations in circuits of the retina”, Neuron, 65, 150–164.
(обратно)12
«Транспортная среда Эймса», спустя почти полвека после изобретения, значится как артикул A1420 в каталоге химической компании Sigma-Aldrich за 2018 г.
(обратно)13
Boycott, B., & Wässle, H. (1999), “Parallel processing in the mammalian retina: The Proctor Lecture”, Investigative Ophthalmology and Visual Science, 40, 1313–1327.
(обратно)14
Энрика исходила из предположения, что так называемая обогащенная среда полезна не только для зрения, но и для развития различных видов мышиного интеллекта. Убедившись в благотворном эффекте такой среды, она провела серию экспериментов, чтобы проверить каждый ее компонент в отдельности. Barone, I., Novelli, E., and Strettoi, E. (2014), “Long-term preservation of cone photoreceptors and visual acuity in rd10 mutant mice exposed to continuous environmental enrichment”, Molecular Vision, 20, 1545–1556.
(обратно)15
Совершенно естественно, что Элио Равиола стал мастером метода Гольджи: он учился на факультете анатомии в Падуанском университете, который в XIX в. окончил Камилло Гольджи.
(обратно)16
За десятилетие, прошедшее с момента публикации этой работы, два идентифицированных Вессле типа биполярных клеток были подразделены еще на два подтипа каждый, поэтому некоторые исследователи считают, что фактически существует 14 типов. Но это отчасти вопрос семантики, и фундаментальное утверждение Вессле остается верным. Wässle, H., Puller, C., Müller, F., and Haverkamp, S. (2009), “Cone contacts, mosaics, and territories of bipolar cells in the mouse retina”, Journal of Neuroscience, 29, 106–117; Helmstaedter, M., Briggman, K. L., Turaga, S. C., Jain, V., Seung, H. S., and Denk, W. (2013), “Connectomic reconstruction of the inner plexiform layer in the mouse retina”, Nature, 500, 168–174.
(обратно)17
Рассказ о Брайане Бойкотте основан на личных воспоминаниях автора, а также на двух прекрасных биографических очерках: Boycott, B. B. (2001), “Brian B. Boycott”, in Squire, L. R., (Ed.), The history of neuroscience in autobiography, San Francisco: Academic Press. Wässle, H. (2002), “Brian Blundell Boycott, 10 December 1924 – 22 April 2000”, Biographical Memoirs of Fellows of the Royal Society, 48, 51–68.
(обратно)18
Для простоты картины я пишу здесь примерно о 30 типах ганглионарных клеток сетчатки, отвечающих за 30 разных репрезентаций. С той же целью я не упоминаю о специфических типах, присущих разным живым видам. На самом деле реальное количество типов ганглионарных клеток, судя по всему, варьируется у разных видов, поскольку их среды обитания предъявляют к их зрению свои специфические требования. У мышей их число может достигать 50. У приматов их предположительно меньше.
Кроме того, разным видам присуща и другая специализация, особенно в распределении клеток по сетчатке. Многие наземные животные имеют «зрительную полосу» – горизонтально вытянутый участок с высокой плотностью нейронов. Это помогает животным-жертвам сканировать горизонт в поисках хищников. У приматов вместо полосы имеется центральная ямка – крошечный участок, на котором плотно упакован особый миниатюрный тип ганглионарных клеток. Именно поэтому человеческое зрение в центре гораздо острее, чем на периферии. Но все сетчатки работают фундаментально одинаковым образом. Все сетчатки выполняют одинаковые операции по нормализации яркости и выделению краев. И все изученные на данный момент сетчатки разбивают изображение на параллельные потоки сигналов, каждый из которых сообщает мозгу об одном аспекте визуальной сцены.
(обратно)19
Stevens, C. F. (1998), “Neuronal diversity: Too many cell types for comfort?”, Current Biology, 8, R708–R710.
(обратно)20
Это только основные места. Небольшие пучки нервных волокон от сетчатки идут в 50 мишеней в головном мозге. Лучше всего изучены претектальные ядра, которые отвечают за направленность взора. Но функции многих других областей пока неизвестны.
(обратно)21
Последние физиологические исследования показывают, что у мышей многие нейроны ЛКТ получают ввод от одного функционального типа ганглионарных клеток сетчатки, тогда как остальные получают смешанный ввод от многих типов. Пока неизвестно, является ли такая организация специфичной для мышей или нет. Roman Roson, M., Bauer, Y., Kotkat, A. H., Berens, P., Euler, T., & Busse, L. (2019), “Mouse dLGN receives functional input from a diverse population of retinal ganglion cells with limited convergence”, Neuron, 102(2), 462–476; Rompani, S. B., Mullner, F. E., Wanner, A., Zhang, C., Roth, C. N., Yonehara, K., and Roska, B. (2017), “Different modes of visual integration in the lateral geniculate nucleus revealed by single-cell-initiated transsynaptic tracing”, Neuron, 93(4), 767–777.
(обратно)22
Точное расположение этих областей не так важно для нашего обсуждения; главное – получить общее понимание. Кроме того, поверхностный контур мозга отличаются значительной вариабельностью – по крайней мере не меньшей, чем у формы носа у людей. Эксперты также расходятся во мнениях относительно их правильного наименования.
(обратно)23
Представление Цао об этом механизме немного отличается от общепринятого. Опираясь на некоторые прямые доказательства, Цао считает, что мозг измеряет значительное число (50) параметров лица – расстояние между глазами и т. д. – и затем объединяет их, чтобы получить отличительную сигнатуру этого конкретного лица. В настоящее время более распространено мнение, что этот механизм менее детерминирован, как описано в главах 10 и 11.
(обратно)24
Карл Лешли (1890–1958), один из первопроходцев в области нейронаук, интересовался, в частности, взаимосвязью структуры мозга и поведения. Он провел длинную серию экспериментов, пытаясь установить связь между областями мозга и памятью – так называемый след памяти, который Ричард Симон назвал энграммой. Лешли был научным руководителем и близким другом Хебба, и его работы легли в основу теории Хебба. Результаты своих исследований Лешли обобщил в классическом эссе: Lashley, K. S. (1950), “In search of the engram”, in Society for Experimental Biology (Ed.), Physiological mechanisms in animal behavior (Society’s Symposium IV) (454–482). Oxford, UK: Academic Press.
(обратно)25
В 1970-е гг. вокруг этой проблемы бушевали жаркие споры. Главные дебаты велись между нативистами – Хьюбелом и Визелем, которые считали, что связи между рецептивными полями запрограммированы природой, и эмпиристами, которые, как и Дональд Хебб, были убеждены, что эти связи в значительной мере зависят от зрительной стимуляции. В одной из статей Хьюбел и Визель утверждали, что избирательность в отношении ориентации у обезьян присутствует от рождения, тем самым опровергая теорию Хебба о том, что обнаружение линий осуществляется сформированными на основе опыта клеточными ансамблями. Однажды я видел краткую – если не сказать грубую – записку Хьюбела Хеббу по этому поводу. Исследования на макаках показали, что у очень молодых животных, еще не получивших значимого зрительного опыта, а также у животных, выросших в среде без линий и контуров, в первичной зрительной коре V1 присутствуют избирательные к ориентации клетки. Все это приуменьшило ценность теории Хебба. Однако в этом случае Хьюбел, который был прав во многих вещах, похоже, ошибался. Вскоре было установлено, что такое предпочтение к ориентации показывает очень небольшое количество клеток, причем не у всех видов макак, и эти клетки имеют гораздо меньшую остроту восприятия по сравнению с нормальными. См.: Espinosa, J. S., and Stryker, M. P. (2012), “Development and plasticity of the primary visual cortex”, Neuron, 75, 230–249.
(обратно)26
Этот рассказ основан на личных воспоминаниях автора, а также на его личных беседах с Дональдом Хеббом, состоявшихся в Честере, Новая Шотландия, в августе 2016 г. Биографические сведения о Хеббе см.: Hebb, D. O. (1980), “D. O. Hebb”, in Lindzey, G. (Ed.) A history of psychology in autobiography, San Francisco; Freeman; Brown, R. E., and Milner, P. M. (2003), “The legacy of Donald O. Hebb: more than the Hebb synapse”, Nature Reviews Neuroscience, 4, 1013–1039.
(обратно)27
Рассказ основан на личных воспоминаниях автора и книге Сейновски об истории искусственного интеллекта: Sejnowski, T. (2018), The deep learning revolution: Artificial intelligence meets human intelligence, Cambridge, MA: MIT Press.
(обратно)28
От CAPTCHA – англ. Completely Automated Public Turing test to tell Computers and Humans Apart – полностью автоматизированный публичный тест Тьюринга для различения компьютеров и людей. – Прим. пер.
(обратно)29
Обычно в качестве капчей используют искаженные изображения букв, которые легко распознаются людьми, но для компьютеров представляют серьезную проблему. В последнее время с этой целью стали также использоваться фотографии – например, набор фотографий автобусов, частично закрытых различными предметами, с инструкцией, содержащей просьбу сосчитать количество автобусов. Между хакерами и разработчиками капчей ведется непрерывная «гонка вооружений».
(обратно)30
В компьютерном зрении признаки, обнаруживаемые на этом этапе, иногда называются априорными. Они основаны на предыдущем опыте распознавания важных классов событий в данной среде. Такие признаки могут просто вводиться в машину или же выделяться посредством машинного обучения. Интересно, что раньше считалось, что всегда лучше задать эти предварительные данные, чтобы уменьшить количество работы, которое нужно проделать компьютеру, – если они нам известны, почему бы не сообщить их алгоритму заранее? Но, как ни странно, впоследствии были получены доказательства того, что гораздо эффективнее позволить алгоритму самому, при помощи машинного обучения, выделить интересующие его признаки.
(обратно)31
В общих чертах: интернейроны сетчатки с большими рецептивными полями – горизонтальные и большие амакриновые клетки – собирают сигналы о яркости с довольно обширного участка сетчатки и затем «вычитают» соответствующий коэффициент яркости из сигнала, который передается от фоторецепторов к ганглионарным клеткам. Есть и другие механизмы как быстрой, так и долговременной адаптации, которые действуют в наружном и внутреннем слоях сетчатки. Например, под воздействием яркого света фоторецепторные клетки снижают свою чувствительность. Амакриновые клетки во внутреннем слое сетчатки непосредственно участвуют в регулировании ответа ганглионарных клеток.
(обратно)32
Описанный здесь базовый механизм был постулирован Хьюбелом и Визелем и непосредственно наблюдался при парной регистрации активности нейронов ЛКТ и зоны V1. Reid, R. C., & Alonso, J. M. (1995), “Specificity of monosynaptic connections from thalamus to visual cortex”, Nature, 378, 281–284.
(обратно)33
Известный феномен – так называемый нейрон Дженнифер Энистон, который был обнаружен при регистрации нейронной активности у пациента во время операции на головном мозге. Удивительно, но этот нейрон возбуждался только в ответ на фотографию этой актрисы и не реагировал на изображения других кинозвезд. Разумеется, лицо Энистон – не единственное лицо, распознаваемое мозгом этого человека; просто ее фотография оказалась у исследователей под рукой. И, по правде говоря, мы не знаем, как нам интерпретировать это открытие. Предположительно «нейрон Дженнифер Энистон» входит в более обширную нейронную сеть, но что это за сеть, у нас пока нет никаких предположений.
(обратно)34
Ископаемые останки мечехвоста Limulus, идентичные по форме современным мечехвостам, были найдены в слоях палеозойской эры. Именно в глазу этого вида ученые впервые обнаружили механизм выделения краев (латеральное торможение). Хотя его «сетчатка» намного проще, чем у млекопитающих, этот механизм работает точно так же: засветка одного фоторецептора вызывает торможение соседних.
(обратно)35
Это мягко сказано. Создатели ИИ не скрывают того, что маленькие дети учатся намного лучше компьютеров. Сегодня часть разработчиков сосредоточилась на создании специализированных систем ИИ, направленных на решение конкретных задач, таких как оценка кредитных рисков. Но другие – мечтатели – поставили перед собой куда более масштабные цели: они хотят создать так называемый общий искусственный интеллект, такой, который окажется в состоянии конкурировать по крайней мере с моим внуком. Подобное изобретение, согласно предположениям, может знаменовать момент технологической сингулярности, когда «верховный алгоритм» запустит нескончаемый цикл самосовершенствований и в конечном итоге захватит мир. Я не смеюсь над сценариями футуристов; на самом деле я рад, что эти мыслители делают свою работу. Но я не волнуюсь на этот счет… пока не волнуюсь. Основная причина в том, что любой верховный алгоритм ограничен – нет, не программным обеспечением, в котором разработчики ИИ демонстрируют впечатляющие успехи, – а оборудованием. Даже знаменитый AlphaZero со всеми его потрясающими возможностями по-прежнему остается узкоспециализированной машиной, умеющей хорошо справляться только со строго определенными задачами и к тому же потребляющей чудовищное количество энергии. Представьте себе машину, способную выполнять все возможные интеллектуальные задачи! Если экстраполировать нынешние параметры, то это будет монстр размером с Ritz, обеспечивать который мегаваттами придется всей Северной Америке.
Хорошо это или плохо, обучающиеся машины в скором времени научатся выполнять довольно сложные задачи. Как оптимист, я верю в лучшее. Кроме того, область ИИ бурлит будоражащими идеями, на которые нам, нейробиологам, стоит обратить внимание.
(обратно)36
В этой главе я привожу пример проблемы, для которой можно предположить хотя бы класс решения. Это проблема связывания мозгом фрагментов визуального образа, и она связана исключительно с физической природой человека. Разные репрезентации существуют в мозге в той или иной физической форме, поэтому можно предположить существование связей между различными признаками или некоего рода нейронного перекрестного сигнала, который говорит мозгу: «Вся эта информация относится к одному и тому же объекту реального мира». Другими словами, это всего лишь проблема поиска связей. Сознание, самость – проблема иного порядка. Содержимое моего сознания известно только мне; это субъективная вещь – да и «вещь» ли это вообще? Может быть, это всего лишь языковой конструкт наподобие парадоксов Зенона? Я думаю, что нет, потому что каждый из нас уверен, что обладает сознанием. Мы не можем отрицать его существование. Но что оно такое на самом деле, остается загадкой. Цитаты: Koch, C. (1982), Consciousness: Confessions of a romantic reductionist, Cambridge, MA: MIT Press.
(обратно)